Дыхательный коэффициент. Источники энергии и пути ее превращения в организме
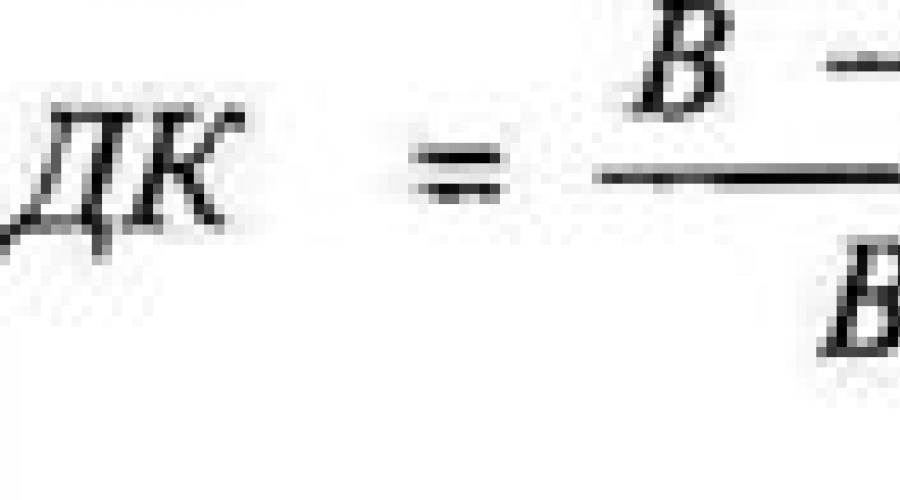
Определение дыхательного коэффициента (ДК) растений.
Дыхательный коэффициент растений - это отношение количества выделенного при дыхании углекислого газа к количеству поглощенного за это же время кислорода.
Величина ДК зависит от химической природы дыхательного субстрата, биологических особенностей растений, условий снабжения кислородом и других причин.
В данной работе рассматривается величина ДК в зависимости от дыхательного субстрата. Если субстратом являются углеводы, то ДК=1; если же субстратом служат более богатые водородом жиры или некоторые белки, ДК меньше 1 и обычно равен 0,3-0,7; когда субстраты - органические кислоты, ДК больше 1.
Ход работы.
Пробирку заполняют до половины проросшими семенами, закрывают пробкой, в которую вставлена изогнутая под прямым углом тонкая стеклянная трубка. Горизонтальное колено трубки должно быть градуировано, или же к нему следует прикрепить полоску миллиметровой бумаги. В трубку ввести каплю вазелинового масла или воды.
Прибор ставят в стакан с ватой (для того, чтобы он не нагрелся от рук). Наблюдают за движением мениска в стеклянной трубке. Если ДК=1, капля остается в трубке неподвижной. Если ДК больше или меньше 1, капля в трубке будет смещаться. Необходимо трижды определить смещение капли за 5 минут и найти среднее значение (А).
А - это разность между объемами поглощаемого Ог и выделяющегося СО2.
Вынуть пробку, проветрить пробирку, в верхнюю часть пробирки вложить диск из фильтровальной бумаги, смоченной 20%-ный раствором КОН. Закрыть пробку, ввести каплю масла, определить передвижение капли за три пятиминутных интервала, вычислить среднее значение (В). Щелочь поглощает выделяемый при дыхании СО2. Передвижение капли теперь соответствует поглощению О2.
Расчеты можно произвести по следующей формуле:
![]()
Оборудование и реактивы:
Широкая плоскодонная пробирка с пробкой, в которую вставлена капиллярная отводная трубка, широкий стакан с ватой, песочные часы на 5 минут, пинцеты, кружочки фильтровальной бумаги, вазелиновое масло, 20%-ный раствор КОН, проросшие семена (пшеницы, подсолнечника, клещевины, фасоли, др.).
Вопросы для повторения:
Термины.
Амилаза - ферменты класса гидролаз, катализирующие гидролиз резервных полисахаридов (крахмала, гликогена). Амилазы обнаружены у животных (сок поджелудочной железы), у высших растений (проросшие семена) и в микроорганизмах. В зависимости от характера действия различают а-амилазы (расщепляют а-1,4-связи в молекуле полисахарида), (З-амилазы (последовательно отщепляют мальтозы от нередуцирующих концов цепей полимера) и глюкоамилазы (расщепляют полисахарид с образованием свободной глюкозы).
Гликолиз - путь Эмбдена-Мейергофа-Парнаеа, ферментативный анаэробный процесс негидролитического распада углеводов до ПВК. Это филогенетически наиболее древний путь, широко распространенный в природе, играет важную роль в обмене веществ живых организмов. Обеспечивает клетку энергией в условиях недостаточного снабжения кислородом.
Дегидрогеназа - ферменты класса оксидоредуктаз, катализирующие реакции отщепления водорода от одного субстрата и переносящие его на другой. Участвуют в процессах катаболизма всех типов питательных веществ. Ко- ферментами дегидрогеназ служат обычно НАД, НАДР, | ФАД, ФМН. Реакции с участием дегидрогеназ лежат в основе биологического окисления, тесно связанного с обеспечением клеток энергией.
Дегидрогеназы анаэробные - двухкомпонентные ферменты, коферментом которых может быть NAD+ и NADP+. При окислении субстрата NAD+ превращаются в восстановленную форму NADH. Анаэробные дегидрогеназы передаюn водород, то есть электроны и протоны, различным промежуточным переносчикам и аэробным дегидрогеназам. Субстратная специфичность фермента зависит от его белковой части. Многие дегидрогеназы содержат ионы двухвалентных металлов, например Zn.
Дегидрогеназы аэробные - двухкомпонентные ферменты, называемые еще флавопротеинами. Кроме белков имеют в составе простетическую группу - рибофлавин (витамин Вг). Донорами электронов аэробных дегидрогеназ служат анаэробные дегидрогеназы, а акцепторами - хиноны, цитохиноны, кислород.
Дыхание - присуще всем органам, тканям и клеткам растений; осуществляется за счет углеводов. Интенсивность дыхания определяется по количеству поглощенного О2 или выделенного ССЬ и зависит от онтогенеза, морфологических особенностей, температуры и т. д.
Дыхателънный коэффициент - отношение объема С02, выделяемого из организма при дыхании, к объему поглощаемого за то же время О2; характеризует особенности газообмена и обмена веществ в живых организмах. Дыхательный коэффициент зависит от химической природы дыхательного субстрат а, содержания СО2 и 02 в атмосфере и др.
Каталаза - фермент класса оксидоредуктаз, катализирует реакцию разложения токсичного для организма перок- сида водорода (Н202) с образованием Н20 и 02. Широко распространенный фермент, содержится в специализированных органоидах - пероксисомах и глиоксисомах. Про- стетическая группа каталазы - гем, в состав которого входит атом железа. Молекулярная масса 250000.
Оксидазы - ферменты класса оксидоредуктаз, катализирующие окислительно-восстановительные реакции, акцепторами водорода в которых служит кислород воздуха. При этом образуется вода или Н202 Коферментом многих оксидаз являются производные витамина В2 - ФАД или ФМН. Оксидазы широко распространены в природе и играют важную роль в катаболизме и детоксикации различных веществ.
Окислительное фосфорилирование - процесс синтеза молекул АТР из АДР и неорганического фосфата за счет энергии окисления молекул органических веществ. Происходит только в живых системах. Открыт данный процесс в 1930 году В. А. Энгельгардом и связан с переносом электронов по электронно-транспортной цепи, встроенной во внутреннюю мембрану митохондрий.
Работа 3. Определение дыхательного коэффициента
Важный показатель химической природы дыхательного субстрата – дыхательный коэффициент (ДК ) – отношение объема выделенного углекислого газа (V (СО 2)) к объему поглощенного кислорода (V (О 2)). При окислении углеводов дыхательный коэффициент равен 1, при окислении жиров (более восстановленных соединений) кислорода поглощается больше, чем выделяется углекислого газа и ДК < 1. При окислении органических кислот (менее восстановленных, чем углеводы соединений) ДК > 1.
Величина ДК зависит и от других причин. В некоторых тканях из-за затрудненного доступа кислорода наряду с аэробным происходит анаэробное дыхание, не сопровождающееся поглощением кислорода, что приводит к повышению значения ДК . Величина дыхательного коэффициента обусловлена также полнотой окисления дыхательного субстрата. Если, кроме конечных продуктов, в тканях накапливаются менее окисленные соединения, то ДК < 1.
Прибор для определения дыхательного коэффициента (рис. 8) состоит из пробирки (рис. 8, а) или другого стеклянного сосуда (рис. 8, б ) с плотно пригнанной пробкой, в которую вставлена измерительная трубка со шкалой из миллиметровой бумаги.
Материалы и оборудование. Прорастающие семена подсолнечника, ячменя, гороха, фасоли, льна, пшеницы, 20 %-й раствор гидроксида натрия, шприц на 2 см 3 , цветная жидкость, чашка Петри, химическая пробирка, U-образно изогнутая трубка, эластичная трубка, пробка с отверстием, пинцет анатомический, полоски фильтровальной бумаги (1,5 5 см), миллиметровая бумага, песочные часы на 3 мин, штатив для пробирок.
Ход работы. В пробирку внесите 2 г прорастающих семян подсолнечника. Плотно закройте пробирку пробкой, соединенной эластичной трубкой с изогнутой U-образно стеклянной трубкой, и введите в конец последней при помощи пипетки небольшую каплю жидкости, создавая внутри прибора замкнутую атмосферу. Во время опыта обязательно поддерживайте постоянную температуру. Для этого поставьте прибор в штатив, избегая тем самым нагревания его руками или дыханием. Определите на сколько делений шкалы продвинется капля внутрь трубки за 3 мин. Для получения точного результата вычислите среднюю величину из трех измерений. Полученная величина выражает разницу между объемом поглощенного при дыхании кислорода и объемом выделенного углекислого газа.
Откройте прибор с семенами и положите в него пинцетом свернутую в кольцо полоску фильтровальной бумаги, предварительно пропитанную раствором NaOH. Снова закройте пробирку, поместите в измерительную трубку новую каплю цветной жидкости и продолжайте измерение скорости ее движения при той же температуре. Новые данные, из которых опять вычислите среднюю величину, выражают объем поглощенного при дыхании кислорода, так как выделившийся углекислый газ поглощается щелочью.
Рассчитайте дыхательный коэффициент по формуле: , где ДК – дыхательный коэффициент; В – объем поглощенного при дыхании кислорода; А – разница между объемом поглощенного при дыхании кислорода и объемом выделенного углекислого газа.
Сравните величины дыхательных коэффициентов предложенных объектов и сделайте вывод о химической природе дыхательных субстратов каждого из объектов.
_________________________________
1 Прибор для наблюдений газообмена при дыхании растений и животных ПГД (учебный): руководство по эксплуатации / под ред. Т.С.Чанова. – М.: Просвещение, 1987. – 8 с.
(например, в листьях и побегах суккулентных растений) и т. д. В зависимости от преимущественного использования тех или иных веществ в процессе дыхания величина дыхательного коэффициента будет изменяться. Когда дыхательным материалом является гексоза, то при полном ее окислении величина, дыхательного коэффициента равняется единице
Увеличение влажности резко усиливает жизнедеятельность и в первую очередь дыхание зерна, сопровождающееся потребностью в кислороде. Вместе с тем запас кислорода в воде очень быстро истощается, например прн замачивании ячменя - за 60-80 мпн, и обеспечение зерна кислородом затруднено . Проникновению кислорода в зерно через зародыш (в начале замачивания) препятствует щиток, а через оболочки впоследствии - большое количество воды в тканях. Диффузия кислорода в воде примерное 10 ООО раз медленнее, чем в газе, кроме того, растворимость его в воде в 40 раз меньше, чем диоксида углерода . Недостаток кислорода в процессе замачивания подтверждается и величиной дыхательного коэффициента, который выше единицы (около 1,07), а через 8 ч от начала замочки равен 1,38, т. е. наблюдается уже анаэробное дыхание.
Фактически же из рис. 60 можно увидеть, что дыхательный коэффициент окисления чайного таннина составляет 0,75, т. е. величину, почти вдвое превышающую теоретически рассчитанную. Интересно отметить, что, по данным Шуберт (1959), дыхательный коэффициент листьев чая в конце составляет 0,7-0,75 факт, свидетельствующий о том, что основным субстратом окислительных процессов в это время служит комплекс катехинов.
Установив величину дыхательного коэффициента прямым определением , делают приближенное вычисление количества превратившихся в организме жиров и углеводов, приняв, что на долю белков приходится обычно около 15% энергии. Для этого можно руководствоваться табл. 16.
Отравление организма сопровождается значительным нарушением обмена веществ. Усиливаются гидролитические процессы , уменьшается содержание в организме гликогена, жиров и липоидов, белковых веществ . Усиление транспирации приводит к значительной потере организмом воды . Уменьшается вес насекомых. Соответственно нарушениям обмена веществ уменьшается дыхательный коэффициент , достигая минимальной величины 0,4-0,5.
Во всяком случае, при фотодинамических процессах потребляется кислород , но это не приводит к образованию СО, так как дыхательный коэффициент (т. е. отношение количества образовавшегося СО2 к количеству поглощенного О2) падает от величины, приблизительно равной единице, до 0,05.
Величина дыхательного коэффициента
Снижение величины дыхательного коэффициента
Интересен вопрос о влиянии света на величину дыхательного коэффициента. Выше уже отмечалось, что выделение СОг листьями на свету у всех видов исследованных растений происходит медленнее, чем у тех же листьев в темноте. Объясняется это тем, что та или иная часть СОг дыхания используется листьями в ходе процессов фотосинтеза. По этой причине ДК листьев на свету всегда ниже, чем тех же листьев в темноте. В особенности отчетливо эти закономерности наблюдаются на суккулентах, в тканях которых, как известно, накапливаются большие количества органических кислот.
Изменения температуры могут резко сказываться на интенсивности поглощения тканями растения кислорода даже и в том случае, если содержание последнего в атмосфере остается неизменным. Наряду с этим температура оказывает мощное влияние не только на общую интенсивность дыхания, но и на соотно-щение между отдельными звеньями этого сложного комплекса процессов. В частности, изменения температуры нередко сильно сказываются на соотнощении между поглощением кислорода и выделением СОг, т. е. на величине дыхательного коэффициента.
Врачи и биологи установили, что при окислении в организме углеводов до воды и углекислого гмза на одну затраченную молекулу кислорода выделяется одна молекула СО2. Таким образом , отношение выделенного СО2 к поглощенному О2 (величина дыхательного коэффициента) равна единице. В случае окисления жиров дыхательный коэффициент равен примерно 0,7. Следовательно, определяя величину дыхательного коэффициента, можно судить, какие вещества преимущественно сгорают в организме. Экспериментально установлено, что при кратковременных, но интенсивных энергия получается за счет окисления углеводов, а при длительных - преимущественно за счет сгорания жиров. Полагают, что переключение организма на окисление жиров связано с истощением резерва углеводов, что обычно наблюдается через 5- 20 мин после начала интенсивной мышечной работы.
Вместо 100 мл начального объема газа при изменившемси давлении в конце опыта имеем 97,68 мл, а 1 мл при этих условних соответствует 0,9768 мл. Последний цифра и ивляется поправочным множителем (К) к первому отсчету объема газа в эвдиометре. Подставляем полученные величины в юрмулу и определяем дыхательный коэффициент
Рис. 61 показывает, что в случае индивидуальных катехинов выделение углекислоты наблюдается лишь через 30 мин. При совместном же окислении этих катехинов выделение углекислоты начинается сразу же и в 3 раза превосходит величину, которую можно рассчитать на основании опытов с отдельными катехинами. Одновременно у смеси катехинов наблюдается и прирост ио-глощения кислорода, но в значительно меньших размерах (-1-45%), чем увеличение выделения углекислоты (- -300%). В результате дыхательный коэффициент возрастает более, чем вдвое.
Макенн и Демусси определяли поправку на дыхание, экспериментируя в темноте Вильштеттер и Штоль доводили поправку на дыхание до ничтожно малой величины , работая на очень сильном свету с высокими концентрациями двуокиси углерода, т. е. в таких условиях , при которых фотосинтез был в 20-30 раз интенсивнее дыхания . В табл. 5 приведены данные из этих работ, а также из некоторых новых исследований, где материалом служили иные типы растений (низшие водоросли). Данные табл. 5 показывают удивительную устойчивость фотосинтетического коэффициента он не зависит от интенсивности света , длительности освещения, температуры, а кислорода и двуокиси углерода. Преобладают значения несколько выше единицы, и отклонения вряд ли превышают предел экспериментальной ошибки . Табл. 5 показывает также, что дыхательный коэффициент
Для соединений, состоящих только из атомов С, О и Н (без перекисных связей), подходящей мерой уровня восстановленности является дыхательный коэффициент (выраженный в виде отношения АСОа/ - ДОд) или еще более удобна обратная ему величина- уровень восстановленности L. Показатель L равен числу молекул кислорода, необходимого для полного сжигания молекулы.
К ресинтезу углеводов, или это чисто окислительный процесс . Если признать правильность теории, доказывающей, что все восстановительные ступени фотосинтеза между комплексами СО) и Н СО должны быть фотохимическими (см. фиг. 20), то темновое превращение яблочной или лимонной кислоты в углеводы кажется невозможным. Уровни восстановленности этих кислот меньше единицы, т. е. они не могут превращаться в углеводы без доступа энергии. Но мы уже рассматривали в главе VH схемы реакций , в которых лишь первая ступень восстановления двуокиси углерода использует световую энергию , а энергия, нужная для последующих ступеней восстановления , доставляется дисмутациями. Таким образом , яблочная и лимонная кислоты могли бы восстанавливаться до углеводов и без помощи света, если часть их будет одновременно окисляться. Подобная энзиматическая дисмутация считается возможной она поддерживается фактом, что дыхательный коэффициент суккулентов во время темнового разрушения кислот часто значительно выше чем 1,33, т. е. величины,. соответствующей сжиганию яблочной кислоты 1212J. В случае чистой дисмутации этот коэффициент должен обратиться в бесконечность. В связи с этими рассуждениями можно привести и другие экспериментальные данные. На стр. 271 указывалось, что в опытах по образованию водорослями крахмала в темноте могли использоваться, как правило, только вещества с i >-1 однако оказалось, что существуют некоторые исключения.
Если листья толстянковых, после того как в них произошло максимальное накопление кислот , оставить в темноте, то их кислотность начинает падать в результате потребления яблочной кислоты с выделением СО2. Это выделение СО2 накладывается на дыхательный обмен , приводя к увеличению дыхательного коэффициента , так что иногда он начинает намного превышать величину 1,33 (это максимальная величина , ожидаемая для полного окисления малата до СО2 и воды). В некоторых, весьма немногочисленных опытах имеются указания на то, что в процессе темнового снижения кислотности происходит некоторое накопление углеводов эти данные служат подтверждением предположения, высказанного много лет назад Беннетом-Кларком согласно этому предположению, в тех случаях, когда наблюдаются очень высокие величины дыхательного коэффициента, происходит потребление части малата в анаболических реакциях . Однако, когда листья, содержащие меченый малат (фиксация С в темноте), подвергали воздействиям , способствующим уменьшению кислотности (к таким воздействиям относится, в частности, повышение температуры), в углеводах листьев обнаруживалось не больше нескольких процентов С. Таким образом , в настоящее время приходится признать , что предположение, согласно которому малат, образовавшийся в процессе ОКТ, превращается в темноте в углеводы в количестве, поддающемся учету, не имеет прямых доказательств если это и возможно, то лишь в исключительных обстоятельствах.
Как уже обсуждалось в предыдущем разделе, растения, у которых протекает ОКТ, обладают выраженной способностью к фиксации СО2. Первым накапливающимся продуктом является малат однако возможно, что изолимонная и лимонная кислоты , накапливающиеся в заметных количествах в листьях таких растений при их развитии, образуются из малата посредством реакций цикла таким образом , в них находится часть углерода, включившегося в листья при темновой фиксации СО2. Такую фиксацию можно легко наблюдать у растений типа толстянковых, так как накопление малата у них происходит быстро и обратимо. В других органах , например в развивающихся листьях, побегах и плодах, кислоты накапливаются относительно медленно и для практических целей необратимо. В этих органах фиксацию СО2, если она происходит, приходится выявлять в таких условиях , когда количество фиксированной СО2 незначительно по сравнению с количеством СО2, выделяющейся в клеточных процессах окисления. Таким образом , в конечном счете можно было бы наблюдать некоторое, возможно, совсем незначительное, понижение величины дыхательного коэффициента по сравнению с той величиной, которую следовало бы ожидать для процессов окисления в органе. Имеются сообщения, что в нескольких случаях наблюдались низкие величины дыхательного коэффициента во время накопления кислот, причем на более ноздних стадиях, когда происходит суммарное расходование кислот, эти величины повышались . Эти наблюдения
Хьюм и др. показали также, что окислительная активность митохондрий, выделенных из яблок (особенно из ткапи кожицы), повышалась на протяжении климактерического периода , причем это повышение начиналось за несколько дней до того, как усиливалось выделение СО2 в целом плоде. (Митохондриальную активность измеряли по поглощению кислорода и выделению углекислоты при добавлении сукцината и малата.) Это наблюдение наряду с тем фактом, что во время климактерического периода несколько возрастало содержание белка, привело Хьюма и его сотрудников к предположению, что в этот период происходит синтез ферментов (пируватдекарбоксилазы и малик-фермента), причем энергия, необходимая для этого синтеза , поступает за счет повышенной митохондриальной активности. Исследователи предположили, далее, что причиной конечного падения интенсивности дыхания до величины, которая остается затем почти постоянной (пока не наступит полный распад ткани), является недостаток кислотного субстрата , необходимого как для цикла Кребса , так и для малик-фермента . Нил и Хьюм показали, что дыхательный коэффициент у дисков из сильно перезревших
Эти длппыс получены Б экспсримбнтзх с кйрпом и серебряным карасем - представителями
Дыхательным коэффициентом называется отношение выделенной при дыхании углекислоты к количеству поглощенного кислорода (СО2/О2). В случае классического дыхания, когда окисляются углеводы СбН^О^ и в качестве конечных продуктов образуются только СО2 и Н2О, дыхательный коэффициент равен единице. Однако так бывает далеко не всегда, в ряде случаев он изменяется в сторону увеличения или уменьшения, почему и считают, что он является показателем продуктивности дыхания. Изменчивость величины дыхательного коэффициента зависит от субстрата дыхания (окисляемого вещества) и от продуктов дыхания (полного или неполного окисления).
При использовании в процессе дыхания вместо углеводов жиров, которые менее окислены, чем углеводы, на их окисление будет использоваться больше кислорода - в таком случае дыхательный коэффициент будет уменьшаться (до величины 0,6 - 0,7). Этим объясняется большая калорийность жиров по сравнению с углеводами.
Если же при дыхании будут окисляться органические кислоты (вещества более окисленные по сравнению с углеводами), то кислорода будет использоваться меньше, чем выделяться углекислоты, и дыхательный коэффициент возрастает до величины больше единицы. Самым высоким (равным 4) он будет при дыхании за счет.щавелевой кислоты, которая окисляется по уравнению
2 С2Н2О4 + 02 4С02 + 2Н20.
Выше было упомянуто, что при полном окислении субстрата (углевода) до углекислого газа и воды дыхательный коэффициент равен единице. Но при неполном окислении и частичном образовании продуктов полураспада часть углерода будет оставаться в растении, не образуя углекислого газа; кислорода будет поглощаться больше, и дыхательный коэффициент опустится до величины меньше единицы.
Таким образом, определяя дыхательный коэффициент, можно получить представление о качественной направленности дыхания, о субстратах и продуктах этого процесса.
Зависимость дыхания от экологических факторов.
Дыхание и температура
Как и другие физиологические процессы, интенсивность дыхания зависит от ряда экологических факторов, причем сильнее и
определеннее всего выражена температурная зависимость. Это обусловлено тем, что из всех физиологических процессов дыхание является наиболее "химическим", ферментативным. Связь же ак- , тивности ферментов с уровнем температуры неоспорима. Дыхание подчиняется правилу Вант-Гоффа и имеет температурный коэффициент (2ю 1,9 - 2,5.
Температурная зависимость дыхания выражается одновершинной кривой (биологической) с тремя кардинальными точками. Точка (зона) минимума различна у разных растений. У холодоустойчивых она определяется температурой замерзания растительной ткани, так что у незамерзающих частей хвойных дыхание обнаруживается при температуре до -25 °С. У теплолюбивых растений точка минимума лежит выше нуля и определяется температурой отмирания растений. Точка (зона) оптимума дыхания лежит в интервале от 25 до 35 °С, т. е. несколько выше, чем оптимум для фотосинтеза. У различных по степени теплолюбивости растений ее положение также несколько изменяется: она лежит выше у теплолюбивых и ниже у холодоустойчивых. Максимальная температура дыхания находится в интервале от 45 до 53 °С.> Эта точка определяется отмиранием клеток и разрушением цитоплазмы, ибо клетка дышит, пока жива. Таким образом, температурная кривая дыхания подобна кривой фотосинтеза, но не повторяет ее. Различие между ними заключается в том, что- кривая дыхания охватывает более широкий температурный диапазон, чем кривая фотосинтеза, а оптимум ее несколько смещен в сторону повышенйой температуры.
Сильное действие на интенсивность дыхания оказывают колебания температуры. Резкие переходы ее от высокой к низкой и обратно значительно усиливают дыхание, что было, установлено* еще В. И. Палладиным в 1899 г.
При колебаниях температуры происходят не только количественные, но и качественные изменения дыхания, т. е. изменение путей окисления органического вещества, однако в настоящее время они исследованьг слабо, поэтому здесь не излагаются.
Отношение объема выделенной двуокиси углерода к объему поглощенного кислорода называется дыхательным коэффициентом.
ДК = СО 2 (л)/О 2 (л)
Дыхательный коэффициент характеризует тип питательных веществ, преимущественно окисляемых в организме на момент его определения. Его рассчитывают, исходя из формул химических окислительных реакций.
Для углеводов :
С 6 Н 12 О 2 + 6О 2 о - 6СО 2 + 6Н 2 О;
ДК = (6 объемов СО 2)/(6 объемов О 2) = 1
Для жиров :
2С 15 Н48,О 6 + 145О 2 о - 102СО 2 + 98Н 2 О;
ДК = (102 объема СО 2)/(145 объемов О 2) = 0,703
Для белков расчет представляет определенную трудность, так как белки в организме окисляются не полностью. Некоторое количество азота в составе мочевины (NH 2) 2 CO 2 выводится из организма с мочой, потом и фекалиями. Поэтому для расчета ДК при окислении белка следует знать количество белка, поступившего с пищей, и количество экскретированных азотсодержащих «шлаков». Установлено, что для окисления углерода и водорода при катаболизме белка и образования 77,5 объема двуокиси углерода необходимо 96,7 объема кислорода. Следовательно, для белков:
ДК = (77,5 объема СО 2)/(96,7 объема О 2) = 0,80
При смешанной пище дыхательный коэффициент составляет 0,8-0,9.
Дыхательный коэффициент при мышечной работе. Главным источником энергии при интенсивной мышечной работе являются углеводы. Поэтому во время работы ДК приближается к единице.
Сразу по окончании работы ДК может резко повыситься. Это явление отражает компенсаторные процессы, направленные на удаление из организма избытка двуокиси углерода, источником которого являются так называемые нелетучие кислоты.
Через некоторое время по завершении работы ДК может резко снизиться по сравнению с нормой. Это связано с уменьшением выделения двуокиси углерода легкими вследствие компенсаторной задержки его буферными системами крови, предотвращающими сдвиг рН в основную сторону.
Примерно через час после завершения работы ДК становится нормальным.
Калорический эквивалент кислорода. Определенному дыхательному коэффициенту соответствует определенный калорический эквивалент кислорода, т.е. количество тепла, которое освобождается при полном окислении 1г питательного вещества (до конечных продуктов) в присутствии 1л кислорода.
Калорический эквивалент кислорода при окислении белков равен 4,8 ккал (20,1 кДж), жиров - 4,7 ккал (19,619 кДж), углеводов - 5,05 ккал (21,2 кДж).
Первоначально газообмен у человека и животных определяли методом Крога в специальных камерах закрытого типа (респираторная камера М.Н. Шатерникова).
В настоящее время полный газовый анализ проводят открытым респираторным методом Дугласа-Xолдейна. Метод основан на сборе выдыхаемого воздуха в специальный приемник (воздухонепроницаемый мешок) с последующим определением общего его количества и содержания в нем кислорода и двуокиси углерода при помощи газоанализаторов.
№ 51 Основной обмен и методы его определения. Условия определения основного обмена и факторы, влияющие на его величину. Специфическое динамическое действие пищи. Закон поверхности М. Рубнера.
Основной обмен - минимальное количество энергии, необходимое для обеспечения нормальной жизнедеятельности в условиях относительного физического и психического покоя. Эта энергия расходуется на процессы клеточного метаболизма, кровообращение, дыхание, выделение, поддержание температуры тела, функционирование жизненно важных нервных центров мозга, постоянную секрецию эндокринных желез.
Печень потребляет 27 % энергии основного обмена, мозг - 19 %, мышцы - 18 %, почки - 10 %, сердце - 7 %, все остальные органы и ткани - 19 %.
Методы определения основного обмена.
Расчет основного обмена по таблицам . Специальные таблицы дают возможность по росту, возрасту и массе тела определить средний уровень основного обмена человека. При сопоставлении этих величин с результатами, полученными при исследовании рабочего обмена с помощью приборов, можно вычислить разницу, эквивалентную затратам энергии для выполнения работы.
Вычисление основного обмена по гемодинамическим показателям (формула Рида). Расчет основан на взаимосвязи между артериальным давлением, частотой пульса и теплопродукцией организма. Формула дает возможность вычислить процент отклонения величины основного обмена от нормы. Допустимым считается отклонение ±10 %.
ПО = 0,75 (ЧСС + ПД 0,74) - 72,
где ПО - процент отклонений; ЧСС - частота сердечных сокращений
(пульс); ПД - пульсовое давление.
Для определения соответствия основного обмена нормативным данным по гемодинамическим показателям существуют специальные номограммы.
Расход энергии в состоянии покоя различными тканями организма неодинаков. Более активно расходуют энергию внутренние органы, менее активно - мышечная ткань. Интенсивность основного обмена в жировой ткани в 3 раза ниже, чем в остальной клеточной массе организма. Люди с низкой массой тела производят больше тепла на 1 кг массы тела, чем с высокой. Если рассчитать энегoвыделение на 1 м 2 поверхности тела, то эта разница почти исчезает. Согласно еще одному правилу Рубнера, основной обмен приблизительно пропорционален поверхности тела для разных видов животных и человека.
Отмечены сезонные колебания величины основного обмена - повышение его весной и снижение зимой. На величину основного обмена влияют предшествующая мышечная работа, состояние желез внутренней секреции.
Условия определения основного обмена.
Любая работа - физическая или умственная, а также прием пищи, колебания температуры окружающей среды и другие внешние и внутренние факторы, изменяющие уровень обменных процессов, влекут за собой увеличение энерготрат.
Поэтому основной обмен определяют в строго контролируемых, искусственно создаваемых условиях: утром, натощак (через 12-14 ч после последнего приема пищи), в положении лежа на спине, при полном расслаблении мышц, в состоянии спокойного бодрствования, в условиях температурного комфорта (18-20 °С). За 3 сут до исследования из рациона исключают белковую пищу. Выражается основной обмен количеством энергозатрат из расчета 1 ккал на 1 кг массы тела в час .
Факторы, определяющие величину основного обмена. Основной обмен зависит от возраста, роста, массы тела, пола человека. Самый интенсивный основной обмен в расчете на 1 кг массы тела отмечается у детей (у новорожденных - 53 ккал/кг в сутки, у детей первого года жизни - 42 ккал/кг). Средние величины основного обмена у взрослых здоровых мужчин составляют 1300-1600 ккал/сут; у женщин эти величины на 10 % ниже. Это связано с тем, что у женщин меньше масса и поверхность тела.
Специфическое динамическое действие пищи - повышение энерготрат организма, обусловленное приемом, перевариванием и усвоением пищи. Специфическое динамическое действие пищи состоит в том, что на переваривание пищи, даже в отсутствии мышечной активности, также расходуется энергия. При этом наибольший расход вызывает переваривание белков. Белки обладают максимально усиливающим действием на обмен веществ, они увеличивают его на 40%, углеводы и жиры увеличивают его всего на 5%. При обычном питании суточный расход на специфическое динамическое действие пищи у взрослого человека составляет около 200 калорий.
Закон поверхности тела Рубнера. Зависимость интенсивности основного обмена от площади поверхности тела была показана немецким физиологом Рубнером для различных животных. Согласно этому правилу, интенсивность основного обмена тесно связана с размерами поверхности тела: у теплокровных организмов, имеющих разные размеры тела, с 1 м 2 поверхности рассеивается одинаковое количество тепла.
Таким образом, закон поверхности тела гласит: энергетические затраты теплокровного организма пропорциональны площади поверхности тела.
С возрастом величина основного обмена неуклонно снижается. Средняя величина основного обмена у здорового человека равна приблизительно 1 ккал/(кг-ч).
№ 52 Рабочий обмен энергии. Энергетические затраты организма при различных видах труда. Методы определения рабочего обмена.
Общий расход энергии человеком зависит от состояния организма и мышечной деятельности.
Мышечная работа сопряжена со значительными затратами энергии (рабочий обмен энергии ), с одной стороны, и увеличением теплопродукции - с другой. У спокойно лежащего человека теплопродукция составляет 35 ккал/(гм 2). Если исследуемый принимает сидячее положение,- на 42 %; в положении стоя - на 70 %, а при спокойной неторопливой ходьбе теплопродукция увеличивается на 180 %. При мышечных нагрузках средней интенсивности КПД работы мышц составляет около 24 %. Из всего количества энергии, расходуемой работающими мышцами, 43 % затрачивается на активацию сокращения, и вся эта энергия переходит в тепло. Только 57 % из общего количества энергии идет на рабочее сокращение.
Разность между энергозатратами при физической нагрузке и энергозатратами основного обмена составляет рабочую прибавку, которая тем больше, чем интенсивнее работа. Рабочая прибавка - это вся остальная энергия, которую тратит организм в течение суток на физическую и умственную активность.
Сумма основного обмена и рабочей прибавки составляет валовый обмен. Сумма валового обмена и специфического динамического действия пищи называется общим обменом.Предельно допустимая по тяжести работа для данного человека, постоянно выполняемая им в течение длительного времени, не должна превышать по энергозатратам уровень основного обмена более чем в 3 раза. При кратковременных нагрузках энергия выделяется за счет окисления углеводов.
При длительных мышечных нагрузках в организме расщепляются преимущественно жиры, обеспечивая 80 % потребной энергии. У тренированных спортсменов энергия мышечных сокращений обеспечивается исключительно за счет окисления жиров. У человека, занимающегося физическим трудом, энергетические затраты возрастают пропорционально интенсивности труда.
По энергетическим затратам все профессии разделены на несколько групп, каждая из которых характеризуется своим суточным расходом энергии.
Коэффициент физической активности. Объективным физическим критерием, определяющим адекватное количество расходования энергии для конкретных профессиональных групп людей, является коэффициент физической активности (отношение общих энерготрат на все виды жизнедеятельности к величине основного обмена, т.е. расходу энергии в состоянии покоя). Величины коэффициента физической активности одинаковы для мужчин и женщин, но в связи с меньшей величиной массы тела у женщин и соответственно основного обмена энерготраты мужчин и женщин в группах с одним и тем же коэффициентом физической активности различны.
Группа I - работники преимущественно умственного труда: научные работники, студенты гуманитарных специальностей. Очень легкая физическая активность; коэффициент физической активности 1,4; расход энергии 1800-2450 ккал/сут.
Группа II - работники, занятые легким физическим трудом: водители трамваев, троллейбусов, работники сферы обслуживания, медицинские сестры, санитарки. Легкая физическая активность; коэффициент физической активности 1,6; расход энергии 2100- 2800 ккал/сут.
Группа III - работники средней тяжести труда: слесари, настройщики, водители автобусов, врачи-хирурги. Средняя физическая активность; коэффициент физической активности 1,9; расход энергии 2500-3300 ккал/сут.
Группа IV - работники тяжелого физического труда: строительные рабочие, металлурги. Высокая физическая активность; коэффициент физической активности 2,2; расход энергии 2850- 3850 ккал/сут.
Группа V - работники особо тяжелого труда, только мужчины: механизаторы, сельскохозяйственные рабочие в посевной и уборочный периоды, горнорабочие, вальщики леса, бетонщики, каменщики, землекопы, грузчики немеханизированного труда, оленеводы и др. Очень высокая физическая активность; коэффициент физической активности 2,5; расход энергии 3750-4200 ккал/сут.
Для каждой группы труда определены средние величины сбалансированной потребности здорового человека в энергии и пищевых веществах, которые несколько различаются для мужчин и женщин.
№ 53 Температура тела человека и ее суточные колебания. Тепловой баланс гомойотермного организма. Температурная схема тела человека. Методы измерения температуры тела человека.
Гомойотермия. В процессе эволюции у высших животных и человека выработались механизмы, способные поддерживать температуру тела на постоянном уровне независимо от температуры окружающей среды. Температура внутренних органов у них колеблется в пределах 36-38 °С, способствуя оптимальному течению метаболических процессов, катализируя большинство ферментативных реакций и влияя в определенных границах на их скорость.
Постоянная температура необходима и для поддержания нормальных физико-химических показателей - вязкости крови, ее поверхностного натяжения, коллоидно-осмотического давления и др. Температура влияет и на процессы возбуждения, скорость и интенсивность сокращения мышц, процессы секреции, всасывания и защитные реакции клеток и тканей.
Гомойотермные организмы выработали регуляторные механизмы, делающие их менее зависимыми от окружающих условий. Они способны избегать перегревания при слишком высокой и переохлаждения при слишком низкой температуре воздуха.
Оптимальная температура тела у человека составляет 37 °С; верхняя летальная температура - 43,4 °С. При более высокой температуре начинается внутриклеточная денатурация белка и необратимая гибель; нижняя летальная температура составляет 24 °С. В экстремальных условиях резких изменений окружающей температуры гомойотермные животные реагируют реакцией стресса (температурный - тепловой или холодовой - стресс). С помощью этих реакций такие животные поддерживают оптимальный уровень температуры тела. Гомойотермия у человека вырабатывается в течение жизни.
Температура тела человека, а также высших животных подвержена более или менее правильным суточным колебаниям даже при одних и тех же условиях питания и физической активности.
Температура тела днем выше, чем ночью, и в течение суток колеблется в пределах 0,5-3 °С, снижаясь до минимального уровня в 3-4 ч утра и достигая максимума к 16-18 ч вечера. Суточный ритм температурной кривой не связан непосредственно со сменой периодов активности и покоя, поскольку он сохраняется и в том случае, если человек постоянно находится в полном покое. Этот ритм поддерживается без каких-либо внешних регулирующих факторов; он присущ самому организму и представляет собой истинно эндогенный ритм.
У женщин выражены месячные циклы колебаний температуры тела. температура повышается после приема пиши (специфическое динамическое действие пищи), при мышечной работе, нервном напряжении.
Температурная схема тела , которая определяется различным уровнем обмена веществ в разных органах. Температура тела в подмышечной впадине - 36,8 °С, на ладонных поверхностях руки - 25-34 °С, в прямой кишке - 37,2-37,5 °С, в ротовой полости - 36,9 °С. Самая низкая температура отмечается в пальцах нижних конечностей, а самая высокая - в печени.
Вместе с тем даже в одном и том же органе существуют значительные температурные градиенты, а ее колебания составляют от 0,2 до 1,2 °С. Так, в печени температура равна 37,8-38 °С, а в мозге - 36,9-37,8 °С. Значительные температурные колебания наблюдаются при мышечной нагрузке. У человека интенсивная мышечная работа приводит к повышению температуры сокращающихся мышц - на 7 °С.
При купании человека в холодной воде температура стопы падает до 16 °С без каких-либо неприятных ощущений.
Индивидуальные особенности температурной схемы тела:
Здоровый человек имеет относительно постоянную температурную схему тела;
Особенности температурной схемы генетически детерминированы, в первую очередь индивидуальной интенсивностью метаболических процессов;
Индивидуальные особенности температурной схемы тела определяются влияниями гуморальных (гормональных) факторов и тонусом вегетативной нервной системы;
Температурная схема тела совершенствуется в процессе воспитания, определяется образом жизни и особенно закаливанием. Вместе с тем она динамична в известных пределах, зависит от особенностей профессии, экологических условий, характера и других факторов.
№ 54 Механизмы теплопродукции. Обмен веществ как источник образования тепла. Роль отдельных органов в теплопродукции и регуляция этого процесса.
Центры теплообразования. В области латерально-дорсального гипоталамуса обнаружены центры теплообразования. Их разрушение приводит к тому, что животные утрачивают способность поддерживать постоянство температуры тела в условиях пониженной температуры окружающей среды. Температура их тела в этих условиях начинает падать, и животные переходят в состояние гипотермии. Электрическое раздражение соответствующих центров гипоталамуса вызывает у животных следующий синдром: 1) сужение поверхностных сосудов кожи. Вазоконстрикция достигается активацией симпатических центров заднего гипоталамуса.; 2) пилоэрекцию- реакция выпрямления волос тела.; 3) мышечную дрожь - увеличивает величину теплопродукции в 4–5 пять раз. Двигательный центр дрожи располагается в дорсомедиальной части заднего гипоталамуса. Он тормозится повышенной внешней температурой и возбуждается при её понижении. Импульсы из центра дрожи вызывают генерализованное повышение мышечного тонуса. Повышенный мышечный тонус приводит к возникновению ритмических рефлексов с мышечных веретён, что и вызывает дрожь; 4) увеличение секреции надпочечников.
Взаимодействие центров терморегуляции. Между центрами теплоотдачи переднего гипоталамуса и центрами теплопродукции заднего гипоталамуса существуют реципрокные взаимоотношения. При усилении активности центров теплопродукции тормозится деятельность центров теплоотдачи и наоборот. При снижении температуры тела включается активность нейронов заднего гипоталамуса; при повышении температуры тела активируются нейроны переднего гипоталамуса.
Механизмы теплопродукции. При снижении температуры окружающей среды эфферентная импульсация от нейронов заднего отдела гипоталамуса распространяется на α-мотонейроны спинного мозга. Эти влияния приводят к сокращению скелетных мышц. При сокращении мышц возрастает гидролиз АТФ. Вследствие этого увеличивается произвольная мышечная активность.
Одновременно при охлаждении возрастает так называемый терморегуляционный тонус мышц. Терморегуляционный тонус представляет своеобразную микровибрацию мышечных волокон. В результате теплопродукция возрастает на 20-45 % от исходного уровня. При более значительном охлаждении терморегуляционный тонус переходит в мышечную холодовую дрожь. Холодовая мышечная дрожь представляет собой непроизвольную ритмическую активность поверхностно расположенных мышц. В результате теплопродукция возрастает в 2-3 раза по сравнению с исходным уровнем.
Механизмы мышечной дрожи связаны с распространением возбуждения из гипоталамуса через покрышку среднего мозга и через красное ядро к α-мотонейронам спинного мозга и от них - к соответствующим мышцам.
Одновременно при охлаждении в скелетных мышцах, печени и буром жире активируются процессы окисления и снижается эффективность окислительного фосфорилирования. За счет этих процессов, так называемого не сократительного термогенеза, теплопродукция может возрасти в 3 раза.
Регуляция несократительного термогенеза осуществляется активацией симпатической нервной системы, гормонами щитовидной железы и мозгового слоя надпочечников.
№ 55 Механизмы теплоотдачи. Способы отдачи тепла организмом. Физиологические механизмы теплоотдачи.
Поддержание температуры тела на оптимальном для метаболизма уровне осуществляется за счет регулирующего влияния ЦНС. За счет нервных и прямых гуморальных влияний, в которых участвует ряд олигопептидов, например бомбезин, в рассматриваемой функциональной системе формируются процессы, направленные на восстановление сформировавшихся изменений температурной схемы тела. Эти процессы включают механизмы теплопродукции и теплоотдачи.
Центры теплоотдачи. В области передних ядер гипоталамуса обнаружены центры теплоотдачи. Разрушение этих структур приводит к тому, что животные утрачивают способность поддерживать постоянство температуры тела в условиях высокой температуры окружающей среды. Температура их тела при этом начинает возрастать, животные переходят в состояние гипертермии, причем гипертермия может развиться даже при комнатной температуре. Раздражение этих структур через вживленные электроды электрическим током вызывает у животных характерный синдром: одышку, расширение поверхностных сосудов кожи, падение температуры тела. Вызванная предварительным охлаждением мышечная дрожь у них прекращается.
Теплоотдачу (физическую терморегуляцию) определяют физические процессы:
Перемещение теплого воздуха с поверхности тела путем контактной или дистантной конвекции;
Теплоизлучение (радиация);
Испарение жидкости с поверхности кожи и верхних дыхательных
Выделение мочи и кала.
Физическая терморегуляция осуществляется следующими путями.
Контактная конвекция - прямой обмен тепла между двумя объектами с разной температурой, находящимися в прямом контакте друг с другом.
Дистантная конвекция - переход тепла в поток воздуха, который движется около поверхности тела и, нагреваясь, заменяется новым, более холодным.
Радиация - отдача тепла путем излучения электромагнитной энергии в
виде инфракрасных лучей.
Регуляция теплоотдачи. Конвекция, теплоизлучение и испарение тепла прямо пропорциональны теплоемкости окружающей среды.
Теплоотдача зависит от объема поверхности тела. Известно, что многие животные на холоде сворачиваются в клубок, занимая меньший объем. Процессы конвекции, излучения и испарения тепла зависят от свойств кожного покрова. Шерстный покров кожи у животных препятствует теплоотдаче.
Сосудистые реакции при перегревании. В основе всех физических процессов теплоотдачи у человека лежат физиологические процессы, связанные с изменением под влиянием окружающей температуры просвета поверхностных сосудов кожи. При действии высокой температуры сосуды расширяются, при действии низкой - суживаются. Эти реакции осуществляются за счет активации вегетативной нервной системы - парасимпатического отдела в первом случае и симпатического - во втором.
В механизмах расширения сосудов кожи принимает участие брадикинин, который продуцируется потовыми железами через холинергические симпатические волокна.
Теплоотдача в водной среде. Процессы теплоотдачи зависят от физических свойств окружающей среды. Наиболее сложно меняются процессы теплоотдачи, так же как и теплопродукции, в водной среде. Прохладная вода обладает наибольшей теплоемкостью. В воде исключается испарение. Одновременно вода оказывает физическое давление на покровы тела, происходит перераспределение массы тела. Температура воды оказывает раздражающее действие на рецепторы кожи и интерорецепторы.
Потоотделение. Наиболее существенным механизмом теплоотдачи является потоотделение. С 1 г пара организм теряет около 600 кал тепла. Потоотделение имеет существенное значение для поддержания оптимального уровня температуры тела в условиях повышенной температуры окружающей среды, особенно в жарких странах. Установлено, что не все люди в равной степени обладают способностью к усиленному потоотделению в условиях повышенной температуры.
№ 56 Функциональная система, поддерживающая оптимальную для метаболизма температуру крови. Характеристика её узловых механизмов.
Функциональная система, определяющая оптимальную для метаболизма температуру тела, объединяет две подсистемы: внутренней эндогенной саморегуляции и целенаправленного поведения. Эндогенные механизмы саморегуляции за счет процессов теплопродукции и тепловыделения определяют поддержание необходимой для метаболизма температуры тела. Функциональная система:
Полезный приспособительный результат
Показатель, ради которого работает данная функциональная система,- температура крови. С одной стороны, она обеспечивает нормальное течение процессов метаболизма, а с другой - сама определяется их интенсивностью.
Для нормального течения метаболических процессов гомойотермные животные, в том числе и человек, вынуждены поддерживать температуру тела на относительно постоянном уровне. Однако это постоянство условно. Температура различных органов подвержена колебаниям, границы которых зависят от времени суток, функционального состояния организма, теплоизоляционных свойств одежды и др.