Взаимосвязь грибов и деревьев. Гриб и водоросль в лишайнике Симбиоз гриба и водоросли вырастает
ЛЕКЦИЯ 8
Отдел Аскомикота – Ascomycota
Это один из наиболее крупных отделов грибов – свыше 32000 видов. Предполагают, что большая часть аскомикота не открыта и общее число видов может быть в 10-20 раз больше. Характерной чертой этой группы является образование после полового процесса аскоспор, которые заключены в сумки или аски. У большинства аскомицетов аск содержит восемь аскоспор и активно выбрасывает споры.
Тело аскомицетов в простейшем случае представлено одиночными почкующимися клетками. У большинства септированный мицелий. Септы, тельца Воронина, транспорт веществ и органелл. У многих видов гифы могут тесно переплетаться (особенно при формировании плодовых тел) и образовывать ложные ткани.
а - стенка гифы, б - септа, в - пора; стрелками показаны тельца Воронина
В состав клеточных стенок входят хитин и глюканы. Хитин составляет меньшую долю полисахаридов клеточной стенки (20-25%), а у дрожжей присутствует лишь в следовых количествах, или вообще отсутствует. Основу клеточной стенки дрожжей составляют глюканы и маннаны (полимеры маннозы). Запасное питательное вещество – гликоген.
Подвижных стадий в цикле развития нет. Бесполое размножение аскомицетов осуществляется конидиями. Коремия, спородохия, ложа, пикнида.
При половом процессе образуются сумки. Половой процесс аскомицетов – гаметангиогамия, иногда соматогамия. Антеридий, аскогон, трихогина, плазмогамия, аскогенные гифы, процесс образования сумок, механизмы освобождения спор, аскокарп или аскома. Гаплоидная, дикариотическая, диплоидная стадии цикла развития.
Половое размножение высших аскомикота

1 – антеридий, аскогон и трихогина;2 - развитие сумок и аскоспор;3 – сумки с 8 спорами.
Отдел аскомикота подразделяется на 5 классов.
Класс археаскомицеты – Archiascomycetes
Эта группа была выделена в результате филогенетического анализа ДНК. Это наиболее древняя линия развития аскомикота. Это очень разнообразная группа по морфологии и образу жизни. Она включает представителей с дрожжеподобной и мицелиальной организацией вегетативного тела. В том числе сюда относится интересный представитель, вызывающий пневмонию (воспаление легких) у людей со сниженным иммунитетом (Pneumocystis ). Плодовые тела не образуют.
Плодовые тела не образуются, сумки развиваются непосредственно на мицелии. У многих настоящий мицелий отсутствует, они представлены одиночными клетками , которые размножаются почкованием или делением.
Порядок Saccharomycetales (устаревшее название – Endomycetales )
На протяжении всего или большей части жизненного цикла дрожжи существуют в виде отдельных клеток. «Дрожжи» - это физиолого-морфологическая категория. Дрожжевые грибы имеются не только у аскомикота, но и у базидиомикота. Предполагают, что дрожжи являются вторично упрощенными формами грибов, жизненной формой, приспособленной к существованию в жидкой среде . Почкование, псевдомицелий, половой процесс. Представители - Saccharomyces cerevisiae , Candida albicans .
Класс плектомицеты – Plectomycetes
Аски образуются в клейстотециях. Это преимущественно почвенные сапротрофы. Многие вызывают деградацию сложных биополимеров, например крахмала и целлюлозы, некоторые способны разрушать кератин.
Порядок эвроциевые – Eurotiales .
Эта группа содержит представителей, анаморфы которых описаны как представители родов Aspergillus, Penicillium. Большинство из них сапротрофы, живущие в почве на растительных остатках. Некоторые развиваются на пищевых продуктах и разных промышленных материалах. Основной тип размножения для многих эвроциевых - образование конидий. У некоторых половое размножение не обнаружено и известны лишь конидиальные стадии– такие грибы вынесены в отдел несовершенных грибов - Deutheromycota.
Класс гименоаскомицеты – Hymenoasco mycetes .
Сумки образуются в гимении или гимениальном слое в аскокарпе или аскоме. Образование аскокарпов происходит после плазмогамии. При этом гаплоидные гифы оплетают развивающиеся аскогенные гифы и сумки, образуя покровную ткань плодового тела. Клейстотеции, перитеции, апотеции. Морфология, способ освобождения спор.
Порядок мучнисторосяные грибы – Erysiphales .
Порядок гипокрейные – Hypocreales .
Характеристика образа жизни, жизненного цикла спорыньи пурпурной (Claviceps purpurea ) и представителей рода Cordyceps.
Порядки Леотиевые и Пецициевые – Leotiales и Pezizales .
Sclerotinia sclerotiorum , пецица фиолетовая – Peziza violacea , сморчок конический – Morchella conica , сморчковая шапочка - Verpa bohemica , строчек обыкновенный - Gyromitra esculenta . Французский трюфель – Tuber melanosporum . Образ жизни трюфелей. Стереотеции.
К ласс асколокулярные или локулоаскомицеты (Loculoascomyces )
Сумки развиваются внутри сплетений гиф (аскостром), которые образуются раньше чем происходит половой процесс. Представитель – возбудитель парши яблони Venturia inaequalis .
ЛЕКЦИЯ 9
Отдел базидиомикота – Basidiomycota (более 30 000 видов).
Базидиомикота характеризуются хорошо развитым многоклеточным мицелием. Подвижные стадии отсутствую, клеточную оболочку образуют глюканы и хитин. Различают первичный и вторичный мицелий (дикариотический). Долипоровые септы. Тип полового процесса – соматогамия. Соотношение гаплоидной и диплоидной фаз. Процесс формирования базидий, пряжки, стеригмы, базидиоспоры. Бесполое размножение, т.е. образование конидий у базидиомицетов встречается редко. Чаще образуются плодовые тела или базидиомы, имеющие гимениальный слой. Парафизы, цистиды. Типы базидий – холобазидия, фрагмо- или гетеробазидия.
КЛАСС ХОЛОБАЗИДИОМИЦЕТЫ – HOLOBASIDIOMYCES .
Класс объединяет грибы с одноклеточными базидиями. Базидии могут формироваться непосредственно на мицелии, быть собраны в гимениальном слое на поверхности плодовых тел, или развиваться внутри плодовых тел.
ГРУППА ПОРЯДКОВ ГИМЕНОМИЦЕТЫ
Это почти половина всех известных базидиомицетов. Базидии развиваются на плодовых телах в гимениальном слое. Плодовые тела могут иметь самую разнообразную форму: у одних они представлены распростертыми по субстрату корочками с гимением на верхней стороне. У других плодовые тела могут быть коралловидными, копытовидными, воронковидными, в виде шляпки на ножке и т.д. Гименофор – гладкий, зубчатый, пластинчатый или трубчатый.
В пределах группы порядков гименомицетов прослеживается эволюция типов плодовых тел и способов размещения на них гименофора. Наиболее примитивными можно считать плоские корковидные плодовые тела с гименофором на верхней поверхности, далее идут плодовые тела с развитой трехмерной структурой и гладким гименофором, покрывающим всю его поверхность, и наконец, наиболее прогрессивны плодовые тела, у которых гименофор поднят над субстратом, находится снизу шляпки и защищен ею, имеет выросты и обеспечивает максимальную продукцию базидиоспор.
Афиллофороидные гименомицеты
Агарикоидные гименомицеты
ГРУППА ПОРЯДКОВ ГАСТЕРОМИЦЕТЫ.
Сейчас некоторые микологи выделяют эту группу из примерно 1100 видов грибов и 14 порядков в самостоятельный класс Gasteromycetes . Основная особенность гастеромицетов состоит в том, что их плодовые тела замкнуты до полного созревания спор. Гимениальный слой находится внутри и к моменту созревания базидиоспор почти всегда разрушается. Высвобождение спор из плодового тела происходит уже после отделения спор от базидий. Гастерокарп, перидий, глеба. Большинство – почвенные сапротрофы, некоторые образуют микоризу с древесными породами. Некоторые виды живут на мертвой древесине. В лесах эти виды принимают участие в разложении валежника.
КЛАСС ГЕТЕРОБАЗИДИОМИЦЕТЫ - HETEROBASIDIOMYCETES
Представители - Rhizoctonia, Auricularia , Tremella .
КЛАСС USTILAGINOMYCETES
Септы простые.
Порядок головневые – Ustilaginales .
КЛАСС UREDINIOMYCETES
Ржавчинные грибы – порядок Uredinales .
ОТДЕЛ ДЕЙТЕРОМИКОТА, ИЛИ НЕСОВЕРШЕННЫЕ ГРИБЫ – DEUTEROMYCOTA
К дейтеромикотам относят грибы, у которых в жизненном цикле половой процесс не наблюдается. В тех случаях, когда половой процесс у тех или иных дейтеромикот обнаруживают, то их переносят в соответствующий таксон другого отдела. Из-за этой особенности все систематические единицы несовершенных грибов являются разнородными по своему происхождению. Виды в них объединяются прежде всего по сходству морфологии и по сходству конидиеобразования. К дейтеромикотам относят около 30 % всех грибов.
Лекция 10.
Лишайники
Изучение срезов лишайников под микроскопом показывает, что внутреннее строение этих организмов неодинаково. Наиболее примитивно устроены некоторые накипные лишайники , у которых клетки водорослей равномерно распределены среди нитей гриба (гифами) по всему слоевищу, как показано на рисунке. Эти лишайники называются гомеомерными.


Талломы более высокоорганизованных лишайников имеют несколько различных слоев клеток, каждый из которых осуществляет определенную функцию. Такие лишайники называют гетеромерными. Снаружи находится защитный коровой слой, состоящий из плотного сплетения грибных гиф и часто окрашенный в серый, коричневый, бурый, желтый, оранжевый и другие цвета. Под верхним коровым слоем размещается слой водорослей. Здесь размещены водорослевые клетки, окруженные тонкими грибными гифами. Чаще всего зона водорослей очень тонка, и поэтому все водоросли равномерно освещаются солнцем через коровой слой. Ниже слоя водорослей лежит сердцевина. Это наиболее толстый слой, определяющий толщину всего лишайникового таллома. Бесцветные грибные тифы сердцевины лежат рыхло, так что между ними остается воздушное пространство. Это обеспечивает свободный доступ внутрь слоевища углекислого газа и кислорода, которые нужны лишайнику для фотосинтеза и дыхания. Снизу таллом, обычно, защищен нижним коровым слоем. Некоторые ученые проводили аналогию между слоевищем листоватых лишайников и листовой пластинкой высших растений . Тесно прилегающие друг к другу грибные гифы корового слоя лишайника напоминают кожицу листа, также выполняющую защитную функцию . Зону водорослей можно сравнить с той тканью листа, где располагаются хлорофиллоносные зеленые клетки и где совершается фотосинтез. Наконец, рыхлая сердцевина слоевища листоватых лишайников сходна с губчатой тканью листа, имеющей воздушные полости . Однако это сходство лишь внешнее, и его причины связаны с аналогичностью функций, которые выполняют части слоевища и лист. У некоторых лишайников на слоевище либо внутри него располагаются особые образования - цефалодии, представляющие собой ассоциацию гриба и синезеленой водоросли. Само слоевище таких лишайников как правило содержит зеленую водоросль. Таким образом, эти лишайники - уже не двух-, а трехкомпонентные организмы: в их состав входят зеленая и синезеленая водоросли, а также гриб. Водорослевый компонент лишайника называется фотобионтом, грибной - микобионтом. Следует предупредить, что термин "фотобионт" появился в литературе о лишайниках сравнительно недавно. Раньше водоросли лишайникового слоевища называли фикобионтом, что в переводе с латинского означает "водорослевый житель", но после того как ученые стали относить синезеленые водоросли к цианобактериям, лихенологи сочли, что более точным будет название "фотобионт", т.е. фотосинтезирующий участник симбиоза. Лишайники, растущие вблизи источников атмосферного загрязнения, если не исчезают совсем, то чаще всего утрачивают свой нарядный, привлекательный вид. На краях лопастей появляется беловатый налет, уменьшается размер слоевищ. На грибных грифах в изобилии появляются бактерии, клетки водорослей уменьшаются в размере, а иногда совсем погибают; бывает, что разрушается весь водорослевый слой таллома. Одним словом, лишайники выглядят больными. Лишайники могут оказать ученым неоценимую услугу как индикаторы загрязнения среды. Еще в 1866 году известный лихенолог В. Нюландер заметил, что в Люксембургском саду Парижа вследствие появления дыма и газов исчезают некоторые виды лишайников. Этим наблюдениям не придавали большого значения , пока развитие промышленности не стало катастрофически сказываться на состоянии лишайников в индустриальных районах. В 1926 году шведский ученый Р. Сернандер опубликовал данные своих лихенологических исследований в Стокгольме. По наличию лишайников он разделил город на три зоны. Центр города с железнодорожными станциями, фабриками и заводами получил название "лишайниковая пустыня" из-за абсолютного отсутствия этих организмов. Вокруг бесплодной зоны лишайниковая флора была бедной, на стволах деревьев и камнях встречались лишь единичные экземпляры. Эта часть города получила название "зона борьбы". Еще дальше, на окраинах, располагалась "нормальная зона" с обычным "набором" лишайников. Лихенологи Н.С. Голубкова и Н.В. Малышева проследили изменение лишайниковой флоры города Казани почти за 100 лет - с 1883 по 1976 год. Сравнив данные за 1976 год с данными за 1913 год, они обнаружили, что по мере развития городского транспорта, промышленности и других видов антропогенного воздействия исчезло 49 видов лишайников. Многочисленные наблюдения в районах промышленных объектов в разных странах показали прямую зависимость между загрязнением атмосферы и уменьшением количества определенных видов лишайников. Были составлены специальные шкалы, отражающие эту зависимость. Пользуясь одной из таких шкал, 15000 английских школьников всего за один 1971 год исследовали распространение лишайников на всей территории Великобритании и сделали карту загрязненности атмосферы.
Симбиоз гриба и водоросли
Страница 3
Цианеллы и свободноживущие сине-зеленые водоросли по своей тонкой организации ничем не отличаются друг от друга. Примечательно, что в цианеллах отсутствуют включения запасных питательных веществ, представленных различными метаболическими гранулами. По всей видимости, надобность в этом отпадает, поскольку цианеллы получают необходимые им вещества прямо из клетки хозяина. В то же время цианеллы поставляют в клетки хозяина некоторые продукты, которые вырабатываются ими в процессе фотосинтеза. Об этом свидетельствует присутствие в цитоплазме бесцветных клеток организма-хозяина крахмальных зерен. Явление это весьма необычное, поскольку у всех хлорофиллоносных зеленых растений единственным местом локализации крахмальных зерен является пластида (хлоропласт). В условиях симбиоза его участники достигают, вероятно, максимальной специализации, в силу которой симбиотирующие сине-зеленые водоросли принимают на себя функции хлоропластов, но ими не становятся. В пользу последнего свидетельствует существенная разница в организации цианелл и пластид. У клеток бесцветного симбионта глаукоцистиса утрачивается способность к самостоятельному образованию крахмала, который образуется там, очевидно, при непосредственном участии цианелл.
Изучение с помощью электронного микроскопа цианелл, входящих в состав глаукоцистиса, выявило у них сильную степень редукции клеточной оболочки . Она сохраняется здесь в виде едва заметного контура, который можно обнаружить лишь при условии высокого качества фиксации и обработки материала. Более тщательное исследование цианелл показало, что их окружает лишь тонкая (100 ^ 10.4) мембрана, называемая плазмалеммой. Такая степень редукции клеточного покрова - уникальное явление среди вступающих в симбиоз сине-зеленых водорослей.
Из приведенной характеристики цианелл видно, что они представляют собой не что иное, как клетки сине-зеленых водорослей, лишенных запасных веществ и клеточных оболочек.
Деление цианелл, как и клеток свободноживущих сине-зеленых водорослей, осуществляется путем перетяжки пополам. Оно автономно и не приурочено к периоду размножения клетки-хозяина. В каждую его дочернюю клетку обычно попадает по несколько цианелл. Таким образом обеспечивается непрерывность симбиоза. В отличие от органелл распределение цнанелл между дочерними клетками хозяина носит случайный характер, поэтому их число там сильно варьирует. Не вызывает никакого сомнения, что само деление и характер расхождения цианелл по дочерним клеткам регулируется не хозяином, что было бы вполне естественно, если бы они превратились в органеллы, а самими цианеллами, сохранившими все свойства клеток. Однако даже в условиях такого высокоразвитого симбиоза, примером которого является глаукоцистис, оба партнера все же сохраняют своп индивидуальные черты и автономность. Об этом свидетельствует их способность к раздельному существованию вне клеток хозяина. В специально подобранной питательной среде изолированные симбионты ведут себя как самостоятельные организмы. Они там не только успешно растут и развиваются, но II размножаются.
Вопрос взаимоотношения гриба и водоросли в слоевище лишайника занимал умы ученых еще в конце прошлого столетия, да и в паше время продолжает волновать лихенологов. Со дня открытия С. Швендснера прошло более 100 лет. За этот период появилось не менее десятка теорий, пытающихся объяснить отношения между грибом и водорослью, однако среди них нет ни одной общепризнанной и окончательно доказанной. С. Ш в е н д е не р, обнаружив, что лишайник состоит из гриба и водоросли, предположил, что гриб в слоевище паразитирует на водоросли. Однако он ошибочно отвел грибу роль хозяина, а водоросли - раба.
Наибольшее распространение среди ученых того времени получила теория мутуалистического симбиоза. Сторонники этой теории считали, что в слоевище лишайника гриб и водоросль находятся во взаимовыгодном симбиозе : водоросль “снабжает” гриб органическими веществами, а гриб “защищает” водоросль от чрезмерного нагревания и освещения и “обеспечивает” ее водой и неорганическими солями. Однако в 1873 г. этой идеалистической теории был нанесен удар. Известный французский исследователь Е.Борн е, изучая анатомическое строение слоевища лишайников, обнаружил внутри водорослевых клеток грибные отростки - гаустории, всасывающие органы гриба. Это позволяло думать, что гриб использует содержимое клеток водорослей, т. е. ведет себя как паразит.
За прошедшие со времен Борне 100 лет в слоевище лишайников было открыто и описано много различных форм абсорбционных, или всасывающих, гиф гриба. Эти гифы плотно прижимаются к клетке водоросли или проникают в нее и служат, как предполагают, для передачи веществ, которые образуют водоросли в результате своей жизнедеятельности, грибному компоненту.
Гриб должен использовать лишь часть водорослей, оставляя резерв - здоровые и нормальные водоросли, содержимым которых он мог бы питаться.
Учеными были замечены любопытные защитные реакции со стороны лишайниковых водорослей. Например, одновременно с проникновением гаустория в клетку водоросли эта клетка делилась. При этом плоскость деления, как правило, проходила как раз через участок, занятый гаусторием, а образовавшиеся в результате этого процесса дочерние клетки были свободны от гаусториев. Было замечено также, что обычно гриб поражает водоросли, уже достигшие определенной стадии зрелости. В молодых растущих водорослях происходит энергичное отложение веществ в оболочке клетки и быстрое ее утолщение. Эта толстая оболочка клетки фикобионта препятствует проникновению абсорбционных органов гриба. Однако большей частью защитная реакция водорослей против активности грибного компонента очень слаба.
Как известно, растения, обитающие на других организмах, но питающиеся самостоятельно, называют э п и ф и т а м и. К эпифитам относится и большая группа водорослей. Особенно часто водоросли эпифитируют на подводных растениях и водоплавающих животных, иногда покрывая их плотным налетом (рис. 46). При эпифитировании между участниками устанавливаются очень непрочные и кратковременные взаимосвязи, которые, однако, уже можно рассматривать как симбиотические. Поскольку эпифитирующая водоросль и хозяин оказывают друг на друга довольно слабое влияние , эпифитизм у водорослей принято считать наиболее примитивной формой симбиоза. Его относят даже к разряду “безразличных”. С подобным утверждением полностью согласиться трудно. Эпифиты действительно не причиняют прямого вреда организму, к которому прикрепляются, но косвенный ущерб при этом все же наносится. Хорошо известно, например, что обрастающие водорослями ножки водоплавающих клещиков, паучков и жучков становятся менее подвижными, а растения сильно затеняются расселившимися на них эпифитами и попадают в условия, неблагоприятные для фотосинтеза. С явлением обрастания нередко приходится сталкиваться при разведении аквариумных растений, которые могут сильно угнетаться обитающими на них водорослями.К сожалению, явление эпифитизма с биологической точки зрения изучено крайне слабо. Не исключено, что между эпифитом и его хозяином устанавливаются взаимоотношения гораздо более сложные, чем
мы обычно себе представляем.
Помимо поверхностного прикрепления, водоросли могут жить в тканях других организмов-как внеклеточно (в слизи, межклетниках, редко в оболочках мертвых клеток, так и внутриклеточно (в содержимом живых неповрежденных клеток. Такие водоросли по способу обитания относят к группе растений эндофитов.
Внеклеточные и особенно внутриклеточные эндофиты из числа водорослей по сравнению с эпифитами образуют более сложные симбиозы-эндосимбиозы. Для них характерно наличие более или менее тесных, постоянных и прочных связей между партнерами. Эндосимбиозы можно выявить только с помощью специальных цитологических исследований.
Наиболее многочисленную группу составляют эндосимбиозы одноклеточных зеленых и желто-зеленых водорослей с одноклеточными животными . Эти водоросли носят названия соответственно зоохлорелл и зооксантелл. Из многоклеточных животных зеленые и желто-зеленые водоросли образуют эндосимбиозы с пресноводными губками, гидрами и др. . Сине-зеленые водоросли образуют с протозоа и некоторыми другими организмами своеобразную группу эндосимбиозов, получивших название синцианозов; возникающий при этом морфологический комплекс из двух организмов называют цианомом, а сине-зеленые водоросли в нем -цианеллами .
Сопоставление между собой различных эндосимбиозов позволяет наметить последовательные ступени усложнения морфологического и функционального соподчинения партнеров. Так, некоторые эндосимбиозы существуют очень непро
Эпифитизм сине-зеленой водоросли Sokolovia neumaniae на ножках водного клещика Neumania triangulares:
должительное время, а затем распадаются, что является свидетельством их примитивности. Примером этого может служить слизистая колониальная сине-зеленая водоросль воронихиния (Woronichinia naegeliana). Почти в 50% случаев в слизи, окружающей шаровидные колонии этой водоросли, живут другие сине-зеленые водоросли (Lyngbya endophytica и Synechocystis endobiotica. Они интенсивно размножаются там, хотя имеют чрезвычайно бледную, едва заметную окраску. Это, вероятно, обусловлено появлением у них способности утилизировать уже готовые органические соединения, которые в изобилии образуются при распаде слизи.
Со временем интенсивное разрастание водорослей в слизи воронихинии приводит сначала к подавлению клеток, а затем к дезорганизации и гибели всей колонии, а, следовательно, и симбиоза в целом.
Возникает вопрос: как проникают водоросли в ткани и клетки других организмов? У некоторых организмов имеются для этого специальные приспособления. Так, у мелкого, плавающего в воде папоротника азоллы (Azolla) на нижней стороне листьев располагаются особые полости с узкими выводными отверстиями, через которые выделяется наружу слизь. В этих полостях, независимо от того, в какой географической точке земного шара растет азолла (в Америке, Азии, Африке или Австралии), поселяются колонии строго определенного вида сине-зеленой водоросли - анабены (Anahaena azollae). Со временем полости закрываются и наступает полная изоляция попавших туда водорослей. Попытки заражения азоллы представителями других родов и даже видов сине-зеленых водорослей успеха не имели. Это свидетельствует о том, что в процессе возникновения данного симбиоза между участниками устанавливается довольно специфическая физиологическая взаимозависимость. Этот вывод подтверждается еще тем, что вырабатываемые азоллой азотистые соединения полностью усваиваются эндосимбиотнрующими здесь экземплярами анабены, вследствие чего у них отпадает свойственная свободноживущим представителям этой сине-зеленой водоросли функция фиксации атмосферного азота. В свою очередь, анабена дополнительно снабжает ткани хозяина кислородом и другими продуктами своей жизнедеятельности.
Несмотря на существующую у этих симбионтов специализацию физиологических процессов ни один из них не претерпевает сколько-нибудь существенных изменений в своей организации.
Однако так обстоит дело далеко не у всех эндосимбиозов подобного типа. Эндосимбиотический образ жизни водорослей чаще всего приводит к частичной или полной редукции их клеточных оболочек. Например, у живущих в тканях морской губки аплизиллы (Aplysilla) особей сине-зеленой водоросли из рода афанокапса (Aphanocapsa) редукция клеточной оболочки выражается в уменьшении ее толщины. За счет этого снижаются защитные свойства оболочки, но повышается ее проницаемость. Последнее качество, несомненно, улучшает условия транспорта
Внеклеточный симбиоз
Поперечный разрез колонии сине-зеленой водоросли воронихии (крупные клетки по периметру), в слизи которой поселяются другие сине-зеленые водоросли синехоцистис(мелкие клетки и лингбия (удлиненные клетки)
Ткань ряски в межклетниках которой поселяется зеленая водоросль хлорохитрум.
Плазмодий желто-зеленой водоросли миксохлорис в мертвой водоносной клетке сфагнума.
Внутриклеточный симбиоз.
1. Амёба с клеточками зеленой водоросли зоохлореллы внутри, вверху отдельная клетка зоохлореллы при большом увеличении.
2 Продольный разрез через конец шупальца пресноводной зеленой гидры (Hydra viridis) С клетками зоохлореллы в клетках внутреннего слоя гидры.
3. Часть таллома зеленой водоросли геосифон (Geosiphon) разветвленные нити которой оканчиваются крупными пузырями в протоплазме которых живет сине-зеленая водоросль носток.
веществ между клетками губки и эндосимбиотирующей там водоросли.
Эндосимбиозы, относящиеся к разряду внеклеточных, образуют уже довольно устойчивые функциональные и морфологические комплексы. Эта тенденция еще более усиливается у внутриклеточных эндосимбиозов. Механизм проникновения водорослей внутрь клеток других организмов без их повреждения и нарушения нормальной жизнедеятельности остается пока нераскрытым. Отчасти предпосылки для возникновения внутриклеточных эндосимбиозов могут быть заложены в сохранении у клеток некоторых организмов голозойного типа питания. Из всех известных типов питания голозойный тип считается одним из наиболее древних.
У организмов с голозойным типом питания захватываемая добыча, в числе которой оказываются и водоросли, поступает непосредственно внутрь клетки и там переваривается. Однако отдельным захваченным особям, вероятно, в силу стечения благоприятных обстоятельств иногда удается не только сохраниться внутри клеток хозяина в неповрежденном виде, но и выработать приспособления к новым, необычным условиям жизни и начать там размножаться. В результате между организмами устанавливаются отношения нового типа - симбиотические. Вероятно, именно так проникают экземпляры подвижной одноклеточной водоросли эвглены (Euglena gracilis) в эпителиальные клетки задней кишки личинок некоторых видов стрекоз. Клетки эвглены остаются там зелеными на протяжении всего периода совместной жизни. Они, правда, теряют подвижность, но при этом никогда не инцистируются. Очевидно, таким же способом особи одноклеточной зеленой водоросли картерии (Carteria) поселяются в эпидермальных клетках ресничного червя конволюта (Convoluta roscoffensis). Как выяснилось, клетки картерии под влиянием симбиотического образа жизни хотя и претерпевают весьма существенные изменения (полностью редуцируется оболочка, и клетки оказываются окруженными только тонкой плазматической мембраной - плазмалеммой, исчезает стигма, упрощается внутренняя организация жгутиков), но не прекращают фотосинтезировать. В свою очередь, червь приобретает способность питаться за счет продуктов жизнедеятельности водоросли, которые вырабатываются в процессе фотосинтеза. В частности, он может жить в течение 4-5 недель, не получая никакой пищи извне. Однако, когда процесс фотосинтеза прекращается (например, если опыт проводить в темноте), гибнут и водоросль, и червь. Более того, личинки червя, лишенные клеток водоросли, не в состоянии вести самостоятельное существование. Искусственное их заражение водорослями не удается.
Внутриклеточные эндосимбиозы, несомненно, легче устанавливаются с теми организмами, клетки которых не имеют жесткой оболочки на протяжении всего жизненного цикла или по крайней мере на одной из его стадий. Проникновение симбионта внутрь клеток с жесткими оболочками возможно только при условии их частичного или полного разрушения. Последнее может наступить под действием специфических энзимов, вырабатываемых организмом, вступающим в симбиотические отношения. Наблюдаемая в ряде случаев строгая специализация вступающих в симбиоз организмов, вероятно. объясняется именно этим обстоятельством. К сожалению, все попытки обнаружить хотя бы следы подобного рода энзимов пока успехом не увенчались.
Одни внутриклеточные эндосимбиозы. как это происходит у личинок стрекоз, периодически распадаются и вновь возобновляются: другие - непрерывно поддерживаются из поколения в поколение, так как в этих случаях между участниками устанавливаются прочные и продолжительные связи. Последняя группа эндосимбиозов. очевидно, могла возникнуть вследствие утраты той фазы в жизненном цикле организма-хозяина, которая была благоприятна для проникновения симбионта в его клетки. С этого момента, по-видимому, и начинается тесная совместная жизнь двух организмов. В таких случаях переход к симбиотическому способу существования неизбежно сопровождается рядом адаптационных изменений у обоих организмов. Иногда эти изменения морфологически незначительны и симбионт можно узнать (например, носток у геосифона, рис. 48, 3), а иногда они настолько существенны, что симбиотирующие водоросли невозможно идентифицировать ни с одной из свободноживущих водорослей.
Так, в вакуолях одного из видов ресничной инфузории парамеции (Paramecium bursaria) неизменно присутствует зеленая одноклеточная водоросль. По морфологии и особенностям поведения ее можно лишь условно отнести к протококковой водоросли из рода хлорелла (Chlorella). Установлено, что клетки водоросли делятся независимо от деления парамеции. Каждая из вновь образующихся дочерних клеток (автоспор) водоросли немедленно заключается в особую вакуолю и в таком виде в дальнейшем распределяется между дочерними особями инфузории.
В ряде случаев между симбионтами складываются настолько тесные взаимозависимые отношения, что вне симбиоза они жить уже не могут. Очевидно, они необратимо утрачивают способность самостоятельно вырабатывать целый ряд веществ, которые в готовом виде поступают от симбиотирующих с ними водорослей. Реальность подобного предположения полностью подтвердилась в опытах с гидрой, которая, оказывается, в нужном количестве получает мальтозу из клетки симбиотирующей там зеленой водоросли, систематическую принадлежность которой точно установить так и не удалось.
Иногда нераспадающиеся эндосимбиозы приводят к образованию такого комплекса, симбио-тическая природа которого выявляется с большим трудом. Так случилось с двумя водорослями - цианофорой и глаукоцистисом.
В 1924 г. была описана новая для науки водоросль, названная цианофорой парадоксальной (Cyanophora paradoxa, табл. 5, 7). Позднее детальное изучение этого организма показало, что цианофора представляет собой симбиоз бесцветной одноклеточной водоросли криптомонады (отдел Pyrrophyta) и поселяющейся в ней внутриклеточно сине-зеленой водоросли (цпанеллы) из рода хроококкус (Chroococcus, отдел Cyanophyta). Клетки последней под влиянием симбиотического образа жизни настолько сильно видоизменяются, что теряют свой типичный облик. Это выражается главным образом в сильной редукции клеточной оболочки.
Она уменьшается не только по толщине, но и по числу входящих в ее состав слоев: вместо четырехслойной, обычно характерной для свободноживущих сине-зеленых водорослей, она становится двухслойной.
Еще большим преобразованиям подвергаются цианеллы, входящие в состав глаукоцистиса (Glaucocystis nostochinearum) - очень своеобразной одноклеточной водоросли, описанной в конце прошлого века. Ее систематическое положение долгое время не удавалось точно определить. На основании сине-зеленой окраски ее сначала отнесли к отделу Cyanophyta. В дальнейшем выявление целого ряда признаков, абсолютно несвойственных сине-зеленым водорослям (наличие морфологически оформленного ядра, окрашенных телец, размножение посредством автоспор), позволили отнести этот организм к зеленым водорослям (отдел Chlorophyta). Только в 30-е годы текущего столетия было наконец установлено, что глаукоцистис представляет собой крайне своеобразную форму эндосимбиоза обесцветившейся одноклеточной водоросли, близкой к роду ооцистис (Oocystis), и палочковидной сине-зеленой водоросли, которая претерпела здесь такие сильные преобразования, что установить точно ее систематическую принадлежность не представляется возможным. В равной степени это может быть любой модифицированный представитель из ряда родов одноклеточных палочковидных сине-зеленых водорослей. В симбиозах подобного рода глаукоцистис является пока единственным примером установления столь тесных взаимоотношений между партнерами. Сине-зеленые водоросли (цианеллы) располагаются в клетках глаукоцистиса либо упорядоченно в виде двух групп, либо беспорядочно, случайно.
Цианеллы и свободноживущие сине-зеленые водоросли по своей тонкой организации ничем не отличаются друг от друга. Примечательно, что в цианеллах отсутствуют включения запасных питательных веществ, представленных различными метаболическими гранулами. По всей видимости, надобность в этом отпадает, поскольку цианеллы получают необходимые им вещества прямо из клетки хозяина. В то же время цианеллы поставляют в клетки хозяина некоторые продукты, которые вырабатываются ими в процессе фотосинтеза. Об этом свидетельствует присутствие в цитоплазме бесцветных клеток организма-хозяина крахмальных зерен. Явление это весьма необычное, поскольку у всех хлорофиллоносных зеленых растений единственным местом локализации крахмальных зерен является пластида (хлоропласт). В условиях симбиоза его участники достигают, вероятно, максимальной специализации, в силу которой симбиотирующие сине-зеленые водоросли принимают на себя функции хлоропластов, но ими не становятся. В пользу последнего свидетельствует существенная разница в организации цианелл и пластид. У клеток бесцветного симбионта глаукоцистиса утрачивается способность к самостоятельному образованию крахмала, который образуется там, очевидно, при непосредственном участии цианелл.
Изучение с помощью электронного микроскопа цианелл, входящих в состав глаукоцистиса, выявило у них сильную степень редукции клеточной оболочки. Она сохраняется здесь в виде едва заметного контура, который можно обнаружить лишь при условии высокого качества фиксации и обработки материала. Более тщательное исследование цианелл показало, что их окружает лишь тонкая (100 ^ 10.4) мембрана, называемая плазмалеммой. Такая степень редукции клеточного покрова - уникальное явление среди вступающих в симбиоз сине-зеленых водорослей.
Из приведенной характеристики цианелл видно, что они представляют собой не что иное, как клетки сине-зеленых водорослей, лишенных запасных веществ и клеточных оболочек.
Деление цианелл, как и клеток свободноживущих сине-зеленых водорослей, осуществляется путем перетяжки пополам. Оно автономно и не приурочено к периоду размножения клетки-хозяина. В каждую его дочернюю клетку обычно попадает по несколько цианелл. Таким образом обеспечивается непрерывность симбиоза. В отличие от органелл распределение цнанелл между дочерними клетками хозяина носит случайный характер, поэтому их число там сильно варьирует. Не вызывает никакого сомнения, что само деление и характер расхождения цианелл по дочерним клеткам регулируется не хозяином, что было бы вполне естественно, если бы они превратились в органеллы, а самими цианеллами, сохранившими все свойства клеток. Однако даже в условиях такого высокоразвитого симбиоза, примером которого является глаукоцистис, оба партнера все же сохраняют своп индивидуальные черты и автономность. Об этом свидетельствует их способность к раздельному существованию вне клеток хозяина. В специально подобранной питательной среде изолированные симбионты ведут себя как самостоятельные организмы. Они там не только успешно растут и развиваются, но II размножаются.
Среди симбиозов, образованных с участием водорослей, наибольший интерес представляет симбиоз водорослей с грибами, известный под названием лишайникового симбиоза.
ВЗАИМООТНОШЕНИЯ ГРИБА И ВОДОРОСЛИ В ТЕЛЕ ЛИШАЙНИКА
За прошедшие со времен Борне 100 лет в слоевище лишайников было открыто и описано много различных форм абсорбционных, или всасывающих, гиф гриба. Эти гифы плотно прижимаются к клетке водоросли или проникают в нее и служат, как предполагают, для передачи веществ, которые образуют водоросли в результате своей жизнедеятельности, грибному компоненту.
гибнет и сам гриб, а значит, перестанет существовать и лишайник.
Гриб должен использовать лишь часть водорослей, оставляя резерв - здоровые и нормальные водоросли, содержимым которых он мог бы питаться.
Учеными были замечены любопытные защитные реакции со стороны лишайниковых водорослей. Например, одновременно с проникновением гаустория в клетку водоросли эта клетка делилась. При этом плоскость деления, как правило, проходила как раз через участок, занятый гаусторием, а образовавшиеся в результате этого процесса дочерние клетки были свободны от гаусториев. Было замечено также, что обычно гриб поражает водоросли, уже достигшие определенной стадии зрелости. В молодых растущих водорослях происходит энергичное отложение веществ в оболочке клетки и быстрое ее утолщение. Эта толстая оболочка клетки фикобионта препятствует проникновению абсорбционных органов гриба. Однако большей частью защитная реакция водорослей против активности грибного компонента очень слаба.
Однако все высказанные по этому поводу точки зрения до сих пор "остаются лишь догадками и большей частью не подтверждены экспериментально: лишайники оказались очень трудным объектом для физиологических исследований. Ученые пока не научились выращивать и поддерживать в живом состоянии слоевище лишайников в искусственных условиях. Тот контакт между грибом и водорослью, который с такой легкостью достигается в природе (достаточно вспомнить многообразие лишайников!), никак не удается воспроизвести в лабораторных условиях. Наоборот, при переносе лишайников в лабораторию этот контакт легко нарушается и растение просто погибает. Время от времени появляются сообщения об удачных опытах выращивания лишайника в условиях лаборатории, но пока эти сообщения единичны и не всегда достоверны.
Одной из причин неудач подобных попыток можно считать чрезвычайно медленный рост лишайников. Лишайники - многолетние растения . Обычно возраст взрослых слоевищ, которые можно увидеть где-нибудь в лесу на стволе деревьев или на почве, составляет не менее 20-50 лет. В северных тундрах возраст некоторых кустистых лишайников рода кладония достигает 300 лет. Слоевище лишайников, имеющих вид корочки, в год дает прирост всего 0,2-0,3 мм.
Кустистые и листоватые лишайники растут несколько быстрее - в год их слоевище увеличивается на 2-3 мм. Поэтому, чтобы вырастить взрослый лишайник в лаборатории, требуется не менее 20 лет, а может быть, и вся жизнь исследователя. Трудно проводить столь долговременный эксперимент!
Вот почему физиологические особенности лишайников, в том числе взаимоотношения компонентов, как правило, изучают на культурах изолированных мико- и фикобионтов. Этот метод очень перспективен, так как позволяет ставить длительные и воспроизводимые опыты. Но, к сожалению, данные, полученные этим методом, не могут полностью отразить те процессы, которые происходят в целом слоевище лишайника.
И тем более мы не вправе считать, что в природе, в естественных условиях , в слоевищах лишайника эти процессы протекают точно так же, как в культурах изолированных симбионтов. Вот почему все теории, пытающиеся объяснить взаимоотношения компонентов лишайников, остаются пока лишь догадками.
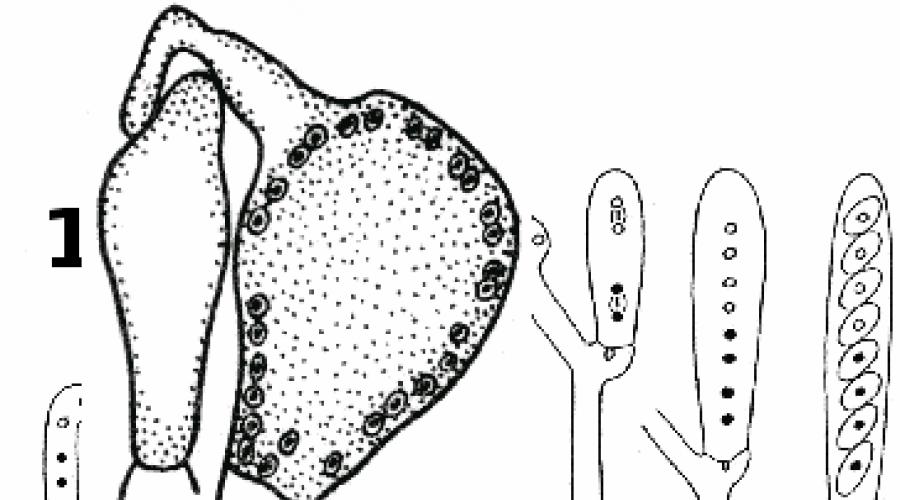
Более успешным оказалось изучение форм контакта между гифами гриба и клетками водорослей в слоевищах лишайников. Как показали исследования с применением электронной микроскопии, в слоевище лишайников можно встретить по крайней мере пять типов контакта между грибными гифами и водорослевыми клетками (рис. 289).
Чаще всего отдельная клетка водоросли и клетка грибной гифы находятся в непосредственном контакте друг с другом. В таком случае гриб образует специальные абсорбционные, всасывающие органы, которые проникают внутрь водорослевой клетки или плотно прижимаются к ее оболочке.
В настоящее время среди абсорбционных органов гриба в слоевище лишайников различают несколько типов: гаустории, импрессории и аппрессории.
Формы контакта между гифами гриба и клетками водорослей в слоевище лищайников.
Гаустории- это боковые выросты гиф гриба, которые прорывают оболочку клетки водоросли и проникают в ее протопласт (рис. 289, 2). Обычно в клетке водоросли развивается одни гаусторий, но иногда их может быть и два. В слоевище лишайника гаустории встречаются в большом количестве и существуют продолжительное время. Было замечено, что в оболочках молодых гаусториев нет отложений целлюлозы, которая могла бы затруднять обмен между клеткой водоросли и гифой гриба. Старые гаустории почти всегда одеты довольно толстым слоем целлюлозы. Различают интрацеллюлярные (внутриклеточные) и интрамембранные (внутриоболочковые) гаустории.
У более высокоорганизованных лишайников образуются только интрамембранные гаустории. Они прорывают оболочку клетки водоросли и достигают ее протопласта, но не углубляются в него, а остаются в оболочке водорослевой клетки (рис. 289, 5). Наибольшее количество интрамембранных гаусториев образуется в слоевище лишайников весной, в начале вегетационного периода. С наступлением осени они далеко отступают от протопласта водоросли.
Другой тип всасывающих органов гриба - импрессории- тоже боковые выросты грибных гиф, но, в отличие от гаусториев, они не прорывают оболочку клетки водоросли, а вдавливают ее внутрь (рис. 289, 6, 7). Импрес copии отмечены у очень многих лишайников, например у широко распространенной пельтигеры (Peltigera).
Интересно, что в слоевищах, произрастающих во влажных местообитаниях, импрессории почти не развиваются, у тех же видов в сухих местообитаниях они образуются в большом количестве. При длительной засухе число импрессориев также увеличивается. Предполагают, что в засушливые периоды и в сухих местообитаниях гриб, чтобы удовлетворить потребности в питании, увеличивает свою всасывающую поверхность за счет увеличения количества и размеров импрессориев.
В отличие от гаусториев и импрессориев, образованных боковыми отростками гифы, аппрессории образуются вершиной грибной гифы. Такая вершина гифы плотно прижимается снаружи к оболочке клетки водоросли, никогда не проникая ни в ее протопласт, ни в ее внутренний слой (рис. 289, 8).
Но в более старых участках слоевища можно найти немало отмерших обесцвеченных клеток - гриб рано или поздно все-таки убивает водоросли. Такой же тип контакта между гифами гриба и клетками водорослей был найден у некоторых слизистых и базидиальных лишайников.
У ряда лишайников, в слоевище которых встречаются нитчатые улотриксовые водоросли, можно наблюдать еще один тип контакта. Как правило, в таком случае нити водорослей бывают целиком покрыты грибными гифами. Причем лишь иногда гифы образуют на поверхности водорослевой нити рыхлую сетку. Чаще же они располагаются очень густо и, срастаясь своими стенками, образуют сплошной чехол. Отдельная лопасть такого лишайника имеет вид тончайшего волоса. Под микроскопом она напоминает полую трубку, стенки которой образованы сросшимися грибными гифами; внутри трубки тянется нить водоросли.
У слизистых лишайников семейства коллемовых (Collemataceae) обычно не наблюдается никакого контакта между грибными гифами и клетками водорослей. Слоевище коллемовых не имеет дифференцированной структуры: нити водоросли носток разбросаны в беспорядке среди грибных гиф по всей толще слоевища (рис. 297, 2). Никаких абсорбционных отростков в клетках водорослей обычно не заметно; гифы гриба и нити сине-зеленой водоросли расположены друг около друга, не вступая в видимый контакт. Предполагают, что в данном случае гриб поглощает органические вещества, ассимилируемые водорослями, прямо из слизи, которая обычно окружает нити ностока. Однако более тщательное изучение этих лишайников показало, что у многих видов коллемы (Collema) в слоевище время от времени образуются специальные абсорбционные гифы, которые тесно прижимаются к одной из клеток водорослевой нити, а через некоторое время можно наблюдать отмирание этой клетки.
Описанные выше формы контакта между гифами микобионта и клетками водорослей, по всей видимости, не исчерпывают всего многообразия способов, с помощью которых гриб и водоросль в слоевище лишайников устанавливают между собой тесную связь. Исследования в этом направлении только начинаются. Можно думать, что дальнейшее изучение тончайших структур лишайникового слоевища с помощью электронного микроскопа не только даст много нового в описании физических контактов между грибным и водорослевым компонентами лишайников, но и откроет новые горизонты в понимании их взаимоотношений
_________________________________________
Пока лишайники являют собой единственный строго доказанный случай возникновения одного совершенно нового организма из двух. Этот факт послужил толчком для поиска синтетических форм в самых различных систематических группах растений и животных. Однако все усилия, приложенные в данном направлении , оказались пока безрезультатными. Тем не менее, предположение о возможности существования синтетических форм организмов оказалось настолько заманчивым, что в биологии появляется новое течение. В отличие от обычных, прочно утвердившихся взглядов биологов на происхождение организмов как на переход от простого к сложному путем дифференциации, зарождается новое представление о возникновении сложного организма из более простых путем синтеза. Некоторые биологи стали рассматривать растительную клетку не как продукт постепенной дифференциации протопласта, а как симбиотический комплекс, синтезированный из нескольких простых организмов . Эти идеи зародились и получили наиболее полное развитие в трудах наших отечественных ученых.
Впервые предположение о важной роли формативного симбиоза (т.е. симбиоза, приводящего к образованию новых форм) в эволюции организмов было высказано академиком А. С. Фаминциным в 1907 г. Развивая дальше эти мысли, К. С. Мережковский в 1909 г. сформулировал гипотезу симбиогенного происхождения организмов и назвал ее “теорией симбиогенезиса”. В дальнейшем она получила широкую известность среди биологов. В 20-е годы ее поддержал и развил дальше известный советский ботаник Б. М. Коз о-П о ля нс кий. В наши дни эти идеи, уже на новом уровне развития биологии, были возрождены американской исследовательницей Сага н-М а р гу л и с в ее гипотезе происхождения эукариотических клеток. В соответствии с этой гипотезой такие клеточные органеллы, как митохондрии, базальные тела жгутиков и пластиды эукариотических клеток, возникли из симбиотирующих в них прокариотических клеток сине-зеленых водорослей и бактерий. В качестве основного довода приводятся некоторые черты сходства в составе, строении и поведении перечисленных органелл и прокариотов. Бесспорно, эти факты заслуживают самого пристального внимания. Однако они недостаточны для обоснования гипотезы симбиогенеза, поскольку черты сходства, как известно, могут появляться у разных по происхождению структур или организмов II вследствие параллелизма в эволюции. Так, система стигма - жгутик у золотистых, желто-зеленых и бурых водорослей по внешнему виду и функциям очень напоминает палочки сетчатки глаза животных, хотя весь процесс их заложения и ход онтогенетического развития говорят о том, что об общности происхождения этих образований не может быть и речи.
Исследование симбиотических организмов в электронном микроскопе показывает, что даже у такого высокоразвитого симбиоза, как глаукоцистис, партнеры сохраняют свои индивидуальные черты и автономность. Анализ симбиозов водорослей с различными организмами выявляет определенную направленность в развитии отношений между партнерами, главным образом по линии максимальной специализации функций и вызванных этим обстоятельством структурных перестроек при сохранении их как самостоятельных организмов; это идет вразрез с положениями гипотезы симбиогенеза. Все это свидетельствует о том, что в настоящее время гипотеза симбиогенеза находится на той стадии разработки, когда логические построения явно преобладают над фактами.
Безусловно, симбиоз может привести к созданию новых организмов, что подтверждается появлением такой своеобразной растительной группы, как лишайники. Отрицать роль симбиоза в эволюции нельзя. И все же очевидно, что это не единственный и не основной путь образования новых форм жизни. С одной стороны, к такому выводу подводит факт существования именно лишайников, поскольку они образуют крайне специализированную и обособленную группу организмов, представляющих слепую ветвь филогенетического развития. С другой стороны, сейчас накапливается большой фактический материал по тонкой организации клетки. Он дает возможность воссоздать картину вероятного обособления и усложнения организации некоторых клеточных органелл у водорослей. Кстати, именно отсутствие подобного рода фактов в свое время как раз и стимулировало зарождение гипотезы симбиогенеза.
Список использованной литературы: “Жизнь растений” (Том 3; Стр72 “Сожительство водорослей с другими организмами”.-Т. В. Седова; Стр385 “Взаимоотношения гриба и водоросли в теле лишайника” Н. С. Голубкова.)
Под ред. профессора М. М. Голлербаха.
Москва “Просвещение” 1977 год
Лекции Н. К Христофоровой “Биотические отношения” (Симбиоз)
Лишайник - это живой организм, образованный симбиозом гриба и водоросли. Водоросли могут быть зелеными водорослями или сине-зелеными водорослями. Сине-зеленые водоросли на самом деле являются бактериями, их называют цианобактерии. Так что лишайник может быть симбиозом 1) гриба и водоросли, или 2) гриба, водоросли и цианобактерии, или 3) гриба и цианобактерии.
Количество различных видов лишайников составляет около 25 тыс. видов. Лишайники встречаются на всех континентах Земли, даже в Антарктиде.
Лишайники встречаются повсеместно, и люди с древних времен их используют в различных целях (как корм для домашних животных, как лекарство и пищу, для окрашивания тканей). Однако люди долгое время не знали, что это за организм. Это стало известно лишь в середине XIX века.
Особенное строение лишайников не дает возможности однозначно определить их какому-то одному царству живого мира. Их можно отнести как к царству растений, так и к царству грибов.
Лишайники растут очень медленно, но живут очень долго. Лишайник может жить сотни и даже тысячи лет.
Тело лишайника представляет собой слоевище. У разных видов лишайников слоевище разное, оно отличается по форме и строению, окраске, размеру. Большинство лишайников имеют слоевище в несколько сантиметров, однако бывают лишайники длиной около метра.
Выделяют три типа лишайников в зависимости от внешнего вида слоевища: накипные, листоватые и кустистые. Накипные лишайники похожи на корку, прилепленную к поверхности, обычно скалы или камня. Листоватый лишайник имеет слоевище в виде пластинок. К поверхности слоевище листоватого лишайника прикреплено толстой короткой ножкой. Кустистый лишайник похож на кустик. Кустик может подниматься над поверхностью или висеть.
Лишайники бывают белым, зеленым, желтым, голубым, серым и другими цветами.
Симбиоз гриба и водоросли в теле лишайника очень тесный, что в результате это дает единый организм. В слоевище переплетаются гифы гриба, между ними располагаются клетки зеленых водорослей или цианобактерий. Эти клетки могут располагаться как одиночно, так и группами.
Таким образом, лишайник сочетает в себе два очень разных организма. Гриб питается гетеротрофно (поглощает готовые органические вещества), а водоросль - автотрофно (синтезирует органические вещества из неорганических). Можно провести аналогию. Микориза - симбиоз между высшими растениями и грибами, а лишайник - симбиоз между низшими растениями и грибами. Однако в лишайнике симбиоз намного теснее. Ведь виды грибов, которые входят в состав лишайников, вообще не могут существовать без водорослей. Хотя большинство водорослей лишайников встречаются в природе отдельно.
Гифы гриба всасывают воду с растворенными минеральными веществами, а водоросли или цианобактерии осуществляют фотосинтез и образуют органические вещества.
Лишайники размножаются участками слоевища и спорами.
Симбиоз водоросли и гриба позволяет жить лишайнику в самых разных и неприспособленных для жизни условиях среды. Лишайники способны расти на скалах, стенах домов, в пустыне и тундре. И, конечно, они повсеместно встречаются в лесах. Однако лишайники очень чувствительны к загрязнениям. Если воздух задымлен, в нем присутствуют вредные газы, то лишайники гибнут. Поэтому лишайники могут служить индикаторами чистоты окружающей среды.
Лишайники первыми заселяют каменистый грунт. В последствие они участвуют в разрушении горных пород, растворяя субстрат. Отмирая, лишайники участвуют в образовании почвы, наряду с другими организмами.
Ягель - это лишайник, который служит кормом для северных оленей. Некоторые виды лишайников съедобны для человека, другие обладают противомикробными свойствами и используются в медицине.
Лишайники - это своеобразная группа живых организмов, тело (слоевище) которых образовано двумя организмами - грибом (микобионт) и водорослью или цианобактерией (фикобионт), находящимися в симбиозе. В составе лишайников обнаружено около 20 тыс. видов грибов и около 26 родов фототрофных организмов. Наиболее часто встречаются зеленые водоросли родов требуксия, трентеполия и цианобактерия носток, являющиеся авто-трофными компонентами примерно у 90% всех видов лишайников.
Рис.1. Лишайники — Arctoparmelia incurva
Симбиотические (мутуалистические) взаимоотношения между компонентами лишайников сводятся к тому, что фикобионт снабжает гриб созданными им в процессе фотосинтеза органическими веществами, а получает от него воду с растворенными минеральными солями. Кроме того, гриб защищает фикобионт от высыхания. Такая комплексная природа лишайников позволяет им получать питание из воздуха, атмосферных осадков, влага росы и туманов, частиц пыли, оседающей на слоевище, из почвы. Поэтому лишайники обладают уникальной способностью существовать в крайне неблагоприятных условиях, часто совершенно непригодных для других организмов, - на голых скалах и камнях, крышах домов, заборах, коре деревьев и др.
Внешнее строение
Тело лишайника представлено слоевищем. Оно очень разнообразно по окраске, размерам, форме и строению. Слоевище может иметь форму тела в виде корочки, листовидной пластинки, трубочек, кустика и небольшого округлого комочка. Некоторые лишайники достигают в длину более метра, но большинство имеют слоевище размером 3-7 см. Они медленно растут – за год увеличиваются на считанные миллиметры, а некоторые – на доли миллиметра. Возраст их слоевища нередко насчитывает несколько сотен и тысяч лет.
Лишайники не имеют типичной зелёной окраски. Окраска лишайников сероватая, зеленовато-серая, светло- или тёмно-бурая, реже жёлтая, оранжевая, белая, чёрная. Окраска обусловлена пигментами, которые находятся в оболочках гиф гриба. Различают пять групп пигментов: зелёные, синие, фиолетовые, красные, коричневые. Цвет лишайников может зависеть также от окраски лишайниковых кислот, которые откладываются в виде кристаллов или зёрен на поверхности гиф.
Живые и отмершие лишайники, скопившаяся на них пыль и песчинки создают не обнажённом грунте тонкий слой почвы, в котором могут закрепиться мхи и другие наземные растения. Разрастаясь, мхи и травы затеняют наземные лишайники, засыпают их отмершими частями своих тел, и лишайники со временем исчезают с этого места. Лишайникам вертикальных поверхностей засыпание не грозит – они разрастаются и разрастаются, впитывая влагу дождей, рос и туманов.
Лишайники встречаются трех типов. Кустистые лишайники похожи на беловатые "кустики" или "косматые бороды", свисающие с ветвей деревьев. Листоватые лишайники представляют собой пластинки на почве или коре деревьев, несколько похожие на сухие листья. Накипные лишайники, похожие на накипь, сплошь покрывают в горах камни и скалы. Размеры лишайников невелики — несколько сантиметров. Окраска разнообразна: желтая, белая, серая, почти черная, красноватая, зеленовато-серая.
Лишайники — симбиотические организмы
На разрезе через любой крупный лишайник хорошо видны клетки водорослей или фотосинтезирующих бактерий, оплетенные гифами гриба. Гриб и водоросль находятся в таком тесном контакте, что возникает как бы единый организм. Водоросль в результате фотосинтеза вырабатывает органические вещества, которые используются грибом. Гриб снабжает водоросль водой и минеральными солями.
Гриб более сильный компонент лишайника, часто угнетающий клетки водорослей. В свободном состоянии гриб лишайника практически не может существовать. Водоросль гораздо меньше зависит от гриба.
Питание лишайника осуществляется обоими симбионтами. Гифы гриба поглощают воду и растворённые в ней минеральные вещества, а водоросль (или цианобактерия), в которой имеется хлорофилл, образует органические вещества (благодаря фотосинтезу).Гифы играют роль корней: они впитывают воду и растворённые в ней минеральные соли. Клетки водорослей образуют органические вещества, выполняют функцию листьев. Воду лишайники впитывают всей поверхностью тела (используют дождевую воду, влагу туманов). Важным компонентом в питании лишайников является азот. Те лишайники, которые в качестве фикобионта имеют зелёные водоросли, получают соединения азота из водных растворов, когда их слоевище пропитывается водой, частично прямо из субстрата. Лишайники, имеющие в качестве фикобионта сине-зелёные водоросли (особенно ностоки), способны фиксировать атмосферный азот.
Размножение лишайников
Размножаются лишайники главным образом кусочками слоевища, а также особыми группами клеток гриба и водоросли, во множестве образующимися внутри его тела. Под давлением их разросшейся массы тело лишайника разрывается, группы клеток разносятся ветром и дождевыми потоками. Кроме того, грибы и водоросли сохранили и свои собственные способы размножения. Грибы образуют споры, водоросли размножаются вегетативным путём. Лишайники размножаются либо спорами, которые образуют микобионт половым или бесполым путём, либо вегетативно – фрагментами слоевища, соредиями и изидиями.
При половом размножении на слоевищах лишайников формируются половые спороношения в виде плодовых тел. Среди плодовых тел у лишайников различают апотеции (открытые плодовые тела в виде дисковидных образований); перитеции (закрытые плодовые тела, имеющие вид маленького кувшина с отверстием наверху); гастеротеции (узкие плодовые тела удлинённой формы). Большинство лишайников (свыше 250 родов) формируют апотеции. В этих плодовых телах споры развиваются внутри сумок (мешковидных образований) или экзогенно, на вершине удлинённо-булавовидных гиф – базидий. Развитие и созревание плодового тела длится 4-10 лет, а затем в течение ряда лет плодовое тело способно продуцировать споры. Спор образуется очень много: так, один апотеций может продуцировать 124 000 спор. Прорастают они не все. Для прорастания нужны условия, прежде всего определённые температура и влажность.
Бесполое спороношение лишайников – конидии, пикноконидии и стилоспоры, возникающие экзогенно на поверхности конидиеносцев. Конидии образуются на конидиеносцах, развивающихся непосредственно на поверхности слоевища, а пикноконидии и стилоспоры – в особых вместилищах пикнидиях. Вегетативное размножение осуществляется кустиками слоевища, а также особыми вегетативными образованиями – соредиями (пылинки – микроскопические клубочки, состоящие из одной или нескольких клеток водорослей, окружённых гифами гриба, образуют мелкозернистую или порошкообразную беловатую, желтоватую массу) и изидиями (маленькие разнообразной формы выросты верхней поверхности слоевища, одного с ней цвета, имеют вид бородавочек, зёрнышек, булавовидных выростов, иногда маленьких листочков).
Экология и значение лишайников
В связи с очень медленным ростом лишайники могут выжить только в местах, не заросших другими растениями, где есть свободные площади для фотосинтеза. На влажных участках они зачастую проигрывают мхам. Кроме того, лишайники проявляют повышенную чувствительность к химическому загрязнению и могут служить его индикаторами. Устойчивости к неблагоприятным условиям способствует невысокая скорость роста, наличие различных способов извлечения и накопления влаги, развитые механизмы защиты. Лишайники, как правило, предъявляют скромные требования к потреблению минеральных веществ, получая их, большей частью, из пыли в воздухе или с дождевой водой, в связи с этим они могут жить на открытых незащищённых поверхностях (камни, кора деревьев, бетон и даже ржавеющий металл). Преимуществом лишайников является терпимость к экстремальным условиям (засухе, высоким и низким температурам (от −47 до +80 градусов по Цельсию, около 200 видов обитают в Антарктике), кислой и щелочной среде, ультрафиолетовому излучению).
В мае 2005 года проводились эксперименты на лишайниках Rhizocarpon geographicum и Xanthoria elegans, показавшие, что эти виды по крайней мере в течение примерно двух недель смогли продержаться вне земной атмосферы, то есть в крайне неблагоприятных условиях. Многие лишайники специфичны к субстрату: одни хорошо развиваются только на щелочных породах, например, известняке или доломите, другие на кислых, не содержащих извести силикатных породах, таких как кварц, гнейс и базальт. Лишайники-эпифиты также предпочитают определённые деревья: выбирают кислую кору хвойных или берёзовых или осно́вную ореховых, клёна или бузины. Ряд лишайников сам выступает в качестве подложки для других лишайников. Нередко формируется типичная последовательность, в которой различные лишайники нарастают друг на друга. Есть виды, которые постоянно живут в воде, например, Verrucaria serpuloides. Лишайники, как и другие организмы, образуют сообщества. Примером лишайниковых ассоциаций является сообщество Cladonio-Pinetum - лишайниковые сосновые леса.
Отдел Лишайники
Строение тела. Лишайники - группа симбиотических организмов, морфологическую основу тела которых образует гриб. В теле лишай-ников сочетаются два компонента: автотрофный - водоросль, или цианобактерия, и гетеротроф-ный - гриб, - образующие еди-ный симбиотический организм.
Интересно, что гриб, составляющий лишайник, в природе вообще не суще-ствует без водоросли, тогда как большинство водорослей, входящих в слоеви-ще лишайника, встречается в свободноживущем состоянии, отдельно от гриба. Физиологически этот тип симбиоза осно-ван на межклеточном обмене между водорослями и гриба-ми.
Симбиоз в биологии лишайников
Гриб питается углеводами водоросли, а водоросли по-лучают от грибов минеральные вещества и воду. Однако симбиоз с грибами приводит к появлению нового биологи-ческого качества, которое выражается у лишайника в его способности размножаться как единый организм.
Вегетативное тело лишайников представлено слоеви-щем, имеющим различную окраску (серую, зеленоватую, буро-коричневую, желтую или почти черную).
Морфологи-чески различают три основных типа слоевища лишайни-ков:
Ø Накипной (корковый). Наиболее низкоорга-низованные - накипные или корковые слоеви-ща, имеющие вид порошковатых, зернистых, бугорчатых налетов, плотно срастающихся с субстратом и не отделяю-щихся от него без значительных повреждений.
Ø Листоватый: пластин-чатое, иногда с волнистым краем, слоевище, горизон-тально расположенное на субстрате (почва, камни, древесина). К субстрату оно прочно прикреплено тол-стой короткой ножкой.
Ø Кустистый. Кустистые лишайники имеют вид кустика, прямостоячего или висячего, сильно разветв-ленного или неразветвленного. Располагаются на поч-ве, а эпифиты - на ветвях деревьев или на скалах. К субстрату прикрепляются небольшими участками сло-евища, а напочвенные - нитевидными ризоидами. меж-ду которыми существуют и переходные формы. Наивысшей организации в своем строении достигают лишайники с кустистым типом слоевища, имеющие вид разветвленного кустика 12-15 см в высоту и срастающиеся с субстратом только основанием.
Лишайник может впитывать влагу всей поверхностью тела, главным образом влагу дождей, росы и туманов. Это позволяет лишайникам поселяться везде, где есть свет. Без света фотосинтез в клетках водоросли не идет, и лишайник погибает.
Большинство лишайников легко переносят высыхание. Фотосинтез и питание у них в это время прекращается, чем и объясняется их незначительный ежегодный прирост.
Размножаются лишайники вегетативно - обломками слоевища или особыми соредиями и изидиями.
Ø Соредии образуются внутри слоевища и состоят из одной или нескольких клеток водоросли, оплетенных гриб-ными гифами. После разрыва коркового слоя они выпада-ют и в виде пыли разносятся ветром.
Ø Изидии являются выростами на поверхности слоевища и тоже состоят из гриба и водоросли. Они обламываются и разносятся ветром, прорастая в новый таллом на субстрате.
Кроме того, наблюдается бесполое размножение при по-мощи спор, самостоятельно образующихся и у водорослей, и у грибов.
Предыдущая18192021222324252627282930313233Следующая
ПОСМОТРЕТЬ ЕЩЕ:
Лишайники - это симбиотические организмы, тело которых (таллом), образованно соединением грибных (микобионт) и водорослевых и/или цианобактериальных (фотобионт) клеток во внешне кажущемся однородным организме.
Лишайники, состоящие из гриба одного вида и цианобактерии (сине-зелёной водоросли) (цианолишайник, например, Peltigera horizontalis) или водоросли (фиколишайник, например, Cetraria islandica) одного вида, называют двухкомпонентными; лишайники, состоящие из гриба одного вида, и двух видов фотобионтов (одной цианобактерии и одной водоросли, но никогда не двух водорослей или двух цианобактерий), называют трёхкомпонентными (например, Stereocaulon alpinum). Водоросли или цианобактерии двухкомпонентных лишайников питаются автотрофно. В трёхкомпонентных лишайниках водоросль питается автотрофно, а цианобактерия, по-видимому, питается гетеротрофно, осуществляя азотфиксацию. Гриб питается гетеротрофно ассимилятами партнера(ов) по симбиозу. Единого мнения о возможности существования свободноживущих форм симбионтов в настоящее время не достигнуто. Имелся опыт выделения всех компонентов лишайников в культуру и последующая реконструкция исходного симбиоза.
Из известных видов грибов в образовании лишайников участвует около 20 %, в основном это аскомицеты (~98 %), остальное базидиомицеты (~0,4 %), некоторые из них, не имея полового размножения, формально относятся к дейтеромицетам. Существуют также актинолишайники, в которых место гриба занимают мицелярные прокариоты актиномицеты. Фотобионт в 85 % представлен зелёной водорослью, встречаются 80 видов из 30 родов, наиболее важным из которых является Trebouxia (входит в состав более чем 70 % видов лишайников). Из цианобактерий (в 10-15 % лишайников) участвуют представители всех крупных групп, кроме Oscillatoriales, наиболее распространён Nostoc. Часты гетероцистные формы Nostoc, Scytonema, Calothrix и Fischerella. В талломе лишайника клетки цианобионта могут структурно и функционально модифицироваться: увеличивается их размер, изменяются форма, уменьшается количество карбоксисом и количество материала оболочек, замедляется рост и деление клеток.
Контакт между компонентами лишайника может быть различен: 1) нет прямого контакта, 2) через поверхности, 3) гриб посредством гаусторий проникает в тело водоросли. Во взаимоотношениях компонентов наблюдается тонкий баланс, так, деление клеток фотобионта согласовано с ростом гриба. Микобионт получает от фотобионта питательные вещества, производимые тем в результате фотосинтеза. Гриб же создаёт водоросли более оптимальный микроклимат: защищает её от высыхания, экранирует от ультрафиолетового излучения, обеспечивает жизнь на кислых субстратах (поставляя фосфаты) смягчает действие ряда других неблагоприятных факторов. Из зелёных водорослей поступают многоатомные спирты, такие как рибит, эритрит или сорбит, которые легко усваиваются грибом. Цианобактерии поставляют в гриб в основном глюкозу, а также азотсодержащие вещества, образуемые благодаря осуществляемой ими фиксации азота. Потоки веществ из гриба в фотобионт не обнаружены.
Считается, что мутуализм (взаимовыгодный симбиоз) двух видов живых существ должен формироваться постепенно, в результате долгой коэволюции. Однако эксперименты американских биологов показали, что многие виды грибов и одноклеточных водорослей могут образовывать мутуалистические системы практически мгновенно, без предшествующего периода взаимной адаптации и без каких-либо генетических модификаций. Для этого гриб и водоросль должны оказаться в среде, где они будут друг для друга единственными источниками необходимых веществ, таких как углекислый газ и аммоний. Исследование подтвердило гипотезу «экологического соответствия», согласно которой не все существующие в природе мутуалистические системы следует трактовать как результат длительной предшествующей коэволюции.
Облигатным (обязательным) мутуализмом называют взаимовыгодные отношения между двумя видами, не способными существовать друг без друга. Принято считать, что такие отношения формируются постепенно, в ходе длительной коэволюции и взаимной адаптации, «притирки» организмов друг к другу. Несомненно, во многих случаях так оно и было (см. Н. Проворов, Е. Долгих, 2006. Метаболическая интеграция организмов в системах симбиоза).
Разумеется, не всякий вид способен встроиться в новое окружение. При интродукции происходит своеобразная сортировка, в ходе которой одни пришельцы приживаются на новом месте, а другие погибают. Так или иначе, приходится признать, что целостное и взаимосвязанное сообщество может сформироваться не только за счет идущей миллионы лет коэволюционной «притирки» видов друг к другу, но и за счет подбора из числа случайных мигрантов таких видов, которые удачно дополняют друг друга и хорошо уживаются вместе. Эту идею, известную под названием ecological fitting (что можно приблизительно перевести как «экологическое соответствие» или «экологический подбор»), начиная с 1980-х годов развивает известный американский эколог Дэниел Джензен (Daniel Janzen).
Могут ли облигатно-мутуалистические системы, обычно считающиеся чем-то вроде апофеоза коэволюции, формироваться по такой же схеме, то есть без всякой коэволюции - просто за счет случайного соответствия двух случайно встретившихся видов, которые при определенных условиях оказываются неспособными жить друг без друга? Эксперименты, проведенные биологами из Гарвардского университета (США), позволяют ответить на этот вопрос утвердительно.
Авторы работали с обычными пекарскими почкующимися дрожжами Saccharomyces cerevisiae и не менее обычными одноклеточными водорослями хламидомонадами (Chlamydomonas reinhardtii ). В природе эти виды в мутуалистических отношениях замечены не были. В лаборатории, однако, они вступили в неразрывную связь легко и быстро, без всякой эволюции или генетических модификаций. Для этого оказалось достаточно выращивать дрожжи и хламидомонады без доступа воздуха в среде, где глюкоза является единственным источником углерода, а нитрит калия - единственным источником азота.
Схема мутуалистических взаимоотношений дрожжей и хламидомонад довольно проста (рис. 1). Дрожжи питаются глюкозой и производят углекислый газ, необходимый хламидомонадам для фотосинтеза (использовать содержащуюся в среде глюкозу хламидомонады не умеют). Водоросли, со своей стороны, восстанавливают нитрит, переводя азот в доступную для дрожжей форму (аммоний). Таким образом, дрожжи обеспечивают хламидомонады углеродом, а хламидомонады снабжают дрожжи азотом. В таких условиях ни один из видов не может расти без другого. Это и есть облигатный мутуализм.
Авторы убедились, что мутуалистическая система благополучно растет в широком диапазоне концентраций глюкозы и нитрита, хотя в одиночку ни один из двух видов в этих условиях не выживает. Только при очень сильном снижении концентрации глюкозы или нитрита рост смешанной культуры прекращается.
Если раскупорить систему, то есть предоставить ей доступ к атмосферному CO 2 , получается сообщество, в котором только один из участников (дрожжи) не может жить без другого, тогда как второй участник (хламидомонады) уже не нуждается в первом для выживания. Впрочем, даже в этом случае хламидомонады лучше растут в присутствии дрожжей, чем без них (очевидно, дополнительный CO 2 , выделяемый дрожжами, идет им на пользу). Таким образом, система остается мутуалистической, хотя со стороны водорослей мутуализм уже не облигатный. Ни один из видов не вытесняет другой.
Если добавить в среду аммоний, получается обратная ситуация: теперь дрожжи могут жить без водорослей (и вообще не нуждаются в них), тогда как водоросли по-прежнему не могут жить без дрожжей. Это уже не мутуализм, а комменсализм (нахлебничество со стороны водорослей). В этом случае дрожжи, которые размножаются быстрее водорослей, заполняют всё жизненное пространство, доводя хламидомонады до вымирания. Авторы предполагают, что устойчивость таких асимметричных систем (в которых только один из участников сильно зависит от другого) определяется соотношением скоростей размножения. Если зависимый вид размножается быстрее, чем независимый, то сожительство двух видов может быть устойчивым; в противном случае независимый вид может полностью вытеснить своего напарника.
Авторы провели аналогичные эксперименты с другими видами хламидомонад и грибов-аскомицетов. Оказалось, что почти все виды дрожжей в данных условиях образуют облигатно-мутуалистические взаимоотношения с хламидомонадами. Правда, продуктивность (скорость роста) симбиотических комплексов оказывается разной. От чего она зависит, определить не удалось: авторы не нашли связи ни со склонностью дрожжей к кислородному дыханию или бескислородному метаболизму (брожению), ни с природными местообитаниями дрожжей, ни со скоростью размножения, ни со степенью влияния концентрации нитритов на рост дрожжей. Очевидно, дело в каких-то других особенностях изученных видов.
Одноклеточная водоросль хлорелла отказалась вступать в мутуалистические отношения с дрожжами, потому что она сама умеет питаться глюкозой и в смешанной культуре вытесняет дрожжи. Не стали образовывать облигатно-мутуалистические комплексы с водорослями дрожжи Hansenula polymorpha , потому что они сами умеют использовать нитрит в качестве источника азота. Но все же исследование показало, что самые разные виды аскомицетов и хламидомонад готовы вступить в симбиотические отношения друг с другом, попав в подходящие условия.
Из многоклеточных (точнее, образующих нитчатые гифы) аскомицетов были протестированы два классических лабораторных объекта - Neurospora crassa и Aspergillus nidulans . Оба вида умеют восстанавливать нитрит и потому не образуют облигатно-мутуалистических систем с хламидомонадами. Однако генетически модифицированные штаммы этих грибов, лишенные способности утилизировать нитрит, вступили в симбиоз с водорослями точно так же, как и дрожжи. Как выяснилось, при этом клетки хламидомонад вступают в непосредственный физический контакт с гифами грибов: под микроскопом видны гифы, обвешанные хламидомонадами, как новогодняя елка (рис. 2).
Мутуалистические взаимоотношения хламидомонад с дрожжами, по-видимому, тоже требуют установления физических контактов между клетками. Об этом свидетельствует тот факт, что систематическое встряхивание смешанной культуры дрожжей и водорослей резко замедляет рост симбиотической системы.
При помощи электронного микроскопа авторы обнаружили плотные контакты, образующиеся между клеточными стенками Aspergillus nidulans и Chlamydomonas reinhardtii , причем клеточная стенка водоросли в местах контакта становится тоньше - возможно, под действием ферментов, выделяемых грибом.
Похожие межклеточные контакты характерны для классических грибно-водорослевых симбиотических систем - лишайников. Аскомицеты в ходе своей эволюции много раз вступали в симбиоз с водорослями и цианобактериями, образуя лишайники. Лишайникообразующие группы разбросаны по всему филогенетическому дереву аскомицетов. Это значит, что такие эволюционные события происходили многократно и независимо в разных эволюционных линиях грибов (см. F. Lutzoni et al., 2001. Major fungal lineages are derived from lichen symbiotic ancestors). По-видимому, аскомицеты в целом «предрасположены» (преадаптированы) к формированию мутуалистических комплексов с одноклеточными водорослями. Эксперименты американских ученых, возможно, проливают свет на ранние стадии формирования таких комплексов.
Впрочем, не следует переоценивать сходство полученных в эксперименте мутуалистических систем с лишайниками. Хотя бы потому, что у большинства лишайников только грибной компонент не может жить в одиночку, тогда как фотосинтезирующие компоненты (одноклеточные водоросли и цианобактерии), как правило, могут прекрасно жить и без гриба. То есть лишайники не являются облигатно-мутуалистическими системами. Да и отсутствие доступа к атмосферному CO 2 вряд ли является проблемой, с которой водорослям часто приходится сталкиваться в природе. Главное в обсуждаемой работе - демонстрация общего принципа. Исследование показало, что облигатный мутуализм может сложиться мгновенно, без всякой эволюции - просто за счет того, что изменившиеся условия делают виды взаимозависимыми. Разумеется, для того, чтобы из такого наспех сформированного симбиотического комплекса развилось что-то действительно сложное и высоко интегрированное, вроде лишайника, без миллионов лет коэволюции уже не обойтись.
Попытки разделить лишайник на гриб и водоросль делались давно, но чаще всего заканчивались неудачей: даже если соблюдались условия стерильности, не всегда была уверенность, что полученная культура - именно лишайниковый симбионт, а не внутренний паразит лишайника. Кроме того, опыты, обычно, не удавалось повторить, а ведь воспроизводимость - одно из главных требований, предъявляемых к эксперименту. Но вот в середине XX века была разработана стандартная методика и изолировано несколько десятков лишайниковых грибов (микобионтов) и лишайниковых водорослей (фотобионтов). Большая заслуга в этой работе принадлежит американскому ученому В. Ахмаджяну.
Итак, в лабораториях, в стерильных пробирках и колбах с питательной средой поселились изолированные симбионты лишайников. Имея в распоряжении чистые культуры лишайниковых партнеров, ученые решились на самый дерзкий шаг - синтез лишайника в лабораторных условиях. Первая удача на этом поприще принадлежит Е. Томасу, который в 1939 году в Швейцарии получил из мико- и фотобионтов лишайник кладония крыночковидная с хорошо различимыми плодовыми телами. В отличие от предыдущих исследователей, Томас выполнял синтез в стерильных условиях, что внушает доверие к полученному им результату. К сожалению, его попытки повторить синтез в 800 других опытах не удались.
Любимый объект исследования В. Ахмаджяна, принесший ему всемирную славу в области лишайникового синтеза, - кладония гребешковая. Этот лишайник широко распространен в Северной Америке и получил простонародное название "британские солдаты": его ярко-красные плодовые тела напоминают алые мундиры английских солдат времен войны североамериканских колоний за независимость. Небольшие комочки изолированного микобионта кладонии гребешковой смешивали с фотобионтом, извлеченным из того же лишайника. Смесь помещали на узкие слюдяные пластинки, пропитанные минеральным питательным раствором и закрепленные в закрытых колбах. Внутри колб поддерживали строго контролируемые условия влажности, температуры и освещенности. Важным условием эксперимента было минимальное количество питательных веществ в среде. Как же вели себя лишайниковые партнеры в непосредственной близости друг к другу? Клетки водоросли выделяли особое вещество, которое "приклеивало" к ним гифы гриба, и гифы сразу начинали активно оплетать зеленые клетки. Группы водорослевых клеток скреплялись ветвящимися гифами в первичные чешуйки. Следующим этапом было дальнейшее развитие утолщенных гиф поверх чешуек и выделение ими внеклеточного материала, а в результате - образование верхнего корового слоя. Еще позже дифференцировались водорослевый слой и сердцевина, совсем как в слоевище природного лишайника. Эти опыты были многократно воспроизведены в лаборатории Ахмаджяна и всякий раз приводили к появлению первичного лишайникового слоевища.
В 40-е годы XX века немецкий ученый Ф. Тоблер обнаружил, что для прорастания спор ксантории настенной требуются добавки стимулирующих веществ: экстрактов из древесной коры, водорослей, плодов сливы, некоторых витаминов или других соединений. Было сделано предположение, что в природе прорастание некоторых грибов стимулируется веществами, поступающими из водоросли.
Примечательно, что для возникновения симбиотических отношений оба партнера должны получать умеренное и даже скудное питание, ограниченные влажность и освещение. Оптимальные условия существования гриба и водоросли отнюдь не стимулируют их воссоединение. Более того, известны случаи, когда обильное питание (например, при искусственном удобрении) вило к быстрому росту водорослей в слоевище, нарушению связи между симбионтами и гибели лишайника.
Если рассматривать срезы лишайникового слоевища под микроскопом, видно, что чаще всего водоросль просто соседствует с грибными гифами. Иногда гифы тесно прижимаются к водорослевым клеткам. Наконец, грибные гифы либо их ответвления могут более или менее глубоко проникать внутрь водоросли. Эти выросты называются гаусториями.
Совместное существование накладывает отпечаток и на строение обоих лишайниковых симбионтов. Так, если свободноживущие синезеленые водоросли родов носток, сцитонема и других образуют длинные, иногда ветвящиеся нити, то у тех же водорослей в симбиозе нити либо скручены в плотные клубочки, либо укорочены до единичных клеток. Кроме того, у свободноживущих и лихенизированных синезеленых водорослей отмечают различия в размерах и расположении клеточных структур. Зеленые водоросли также изменяются в симбиотическом состоянии. Это, в первую очередь, касается их размножения. Многие из зеленых водорослей, живя "на свободе", размножаются подвижными тонкостенными клеточками - зооспорами. В слоевище зооспоры, обычно, не образуются. Вместо них появляются апланоспоры - относительно маленькие клетки с толстыми стенками, хорошо приспособленные к засушливым условиям. Из клеточных структур зеленых фотобионтов наибольшим изменениям подвергается оболочка. Она тоньше, чем у тех же водорослей "на воле", и имеет ряд биохимических различий. Очень часто внутри симбиотических клеток наблюдают жироподобные зернышки, которые после изъятия водоросли из слоевища исчезают. Говоря о причинах этих различий, можно предположить, что они связаны с каким-то химическим воздействием грибного соседа водоросли. Сам микобионт также испытывает воздействие водорослевого партнера. Плотные комочки изолированных микобионтов, состоящие из тесно переплетенных гиф, внешне совсем не похожи на лихенизированные грибы. Внутреннее строение гиф тоже различно. Клеточные стенки гиф в симбиотическом состоянии значительно тоньше.
Итак, жизнь в симбиозе побуждает водоросль и гриб менять свой внешний облик и внутреннее строение.
Что же получают сожители друг от друга, какую пользу извлекают из совместного существования? Водоросль снабжает гриб, своего соседа по лишайниковому симбиозу, углеводами, полученными в процессе фотосинтеза. Водоросль, синтезировав тот или иной углевод, быстро и почти целиком отдает его своему грибному "сожителю". Гриб получает от водоросли не только углеводы. Если синезеленый фотобионт фиксирует атмосферный азот, существует быстрый и устойчивый отток образовавшегося аммония к грибному соседу водоросли. Водоросль же, очевидно, просто получает возможность широко расселяться по Земле. По словам Д. Смита, "наиболее частая у лишайников водоросль, требуксия, очень редко живет вне лишайника. Внутри же лишайника она распространена, пожалуй, шире, чем любой род свободноживущих водорослей. Цена за занятие этой ниши - снабжение гриба-хозяина углеводами".
При использовании материалов сайта, необходимо ставить активные ссылки на этот сайт, видимые для пользователей и поисковых роботов.
Грибы и деревья

Лишайники

Грибы и орхидеи
Микотизм

Грибы и фауна



Если вам понравилось, прочитайте и другие наши статьи о симбиозе различных организмов: